 |
|
von Reinhard Junker
Studium Integrale Journal
26. Jahrgang / Heft 1 - April 2019
Seite 11 - 18
|
 |
Zusammenfassung: Wie können Feder- und Flugentstehungshypothesen getestet und ggf. widerlegt werden? In der Praxis wird bei historischen Rekonstruktionen nicht mit Falsifizierung gearbeitet, sondern es wird nach bestätigenden Befunden gesucht. Das kann aber nur funktionieren, wenn alle Antwortoptionen zugelassen werden.
|
|
|
Einführung
Die bisherigen fünf Teile dieser Artikelserie handelten von Bau und Feinbau der Vogelfedern und ihrer Verankerung; Modelle zu ihrer Entstehung wurden erläutert und bewertet, fossile Funde vorgestellt, die Hinweise zur Entstehung von Federn geben könnten, und Flugentstehungstheorien besprochen. Es wurde gezeigt, dass alle vorgeschlagenen Modelle zur Entstehung des Vogelflugs vage sind, kaum die zahlreichen für Federn und Flug erforderlichen Details beachten, große Änderungsschritte beinhalten, die von einer selektierbaren Fortbewegungsweise zur nächsten überwunden werden müssten, und kaum testbar sind.
Bezüglich des letzten Punktes stellt sich die Frage, wie historische Hypothesen (Hypothesen über die erstmalige Entstehung) überhaupt getestet und ggf. falsifiziert werden können. Einige Autoren betrachten auf diesem Gebiet Falsifizierung (Versuche der Widerlegung) als falschen Weg; vielmehr seien in historischen Fragen bestätigende Befunde gefragt. Solche methodischen Fragen werden überraschend selten in der biologischen Fachliteratur behandelt. Im Folgenden sollten zunächst Kriterien vorgestellt werden, anhand derer Feder- und Flugentstehungstheorien getestet werden. Danach wird der Frage nachgegangen, warum „Schöpfung“ als Erklärung eine – gemessen an den vorliegenden Daten – gut begründete Option ist.
|
Ein Kennzeichen naturwissenschaftlicher Hypothesen ist ihre Testbarkeit, also die Möglichkeit, ihre Aussagen anhand empirischer* Daten (Beobachtungen) zu überprüfen. Wie aber kann empirisch geprüft werden, welche Vorgänge in der Natur in der Vergangenheit abgelaufen sind? Singuläre vergangene Ereignisse sind anders zu testen als regelhaft ablaufende Prozesse, die experimentell erforscht werden können. Direkte experimentelle Untersuchungen sind nicht möglich und es handelt sich nicht um gesetzmäßig beschreibbare Prozesse, die durch falsifizierende Befunde widerlegt werden könnten. Tatsächlich werden naturhistorische Rekonstruktionen weniger durch Falsifikationen konkurrierender Hypothesen als vielmehr durch bestätigende Befunde geprüft und es wird abgewogen, welche der konkurrierenden Hypothesen die meisten und überzeugendsten Befunde für sích verbuchen kann.
Dies ist auch der Fall bei der Bewertung von Hypothesen zur Entstehung des Vogelflugs. Hier stehen sich klassischerweise die Cursorial- und die Arborealhypothese gegenüber (Entstehung des Vogelflugs vom Boden oder von Bäumen aus), neuerdings auch Mischhypothesen aus diesen beiden Ansätzen. In der kontroversen Diskussion über diese Konkurrenten werden verschiedene Prüfkriterien angewendet: Stimmigkeit mit Fossilfunden, Stimmigkeit mit der Phylogenie, Stimmigkeit mit Kenntnissen über Evolutionsmechanismen; Plausibilisierungen kleinschrittiger, adaptiver Stadien. Es zeigt sich, dass diese Kriterien entweder nicht als Tests geeignet sind, weil sie zu vage sind und nicht den in Rede stehenden Vorgang testen, oder dass die vorgelegten Modelle den Test nicht bestehen. Häufig wird mit Kritik am Konkurrenten argumentiert, was aber ebenfalls nicht als Test für die bevorzugte Hypothese gelten kann.
Für die Flugentstehungshypothesen werden allerdings nur natürliche, blinde, nichtgeistige Naturprozesse, die gesetzmäßig oder regelhaft beschreibbar sind, sowie zufällige Randbedingungen zugelassen. Das Scheitern der Erklärungsversuche auf dieser Basis und eine Reihe typischer Design-Indizien können zum Anlass genommen werden, den grundlegend andersartigen Erklärungsansatz „Schöpfung“ zuzulassen. Für diesen Ansatz gibt es eine Reihe guter Gründe, die abschließend zusammengefasst werden.
|
|
|
Testbarkeit der Flugentstehungsmodelle
Verschiedene Autoren haben hin und wieder festgestellt, dass Theorien zur Entstehung des Vogelflugs nicht testbar seien. Beispielsweise äußern Burgers & Chiappe 1999, 62), dass Erklärungen der Entstehung des Fluges auf Mutmaßungen beruhen und es unwahrscheinlich sei, dass sie jemals überprüft werden können.1 Diesbezüglich kann man folgende Feststellung geradezu als Offenbarungseid lesen: „Unfortunately, we can never know how the different forms of animal flight began“ (Ostrom 1986, 81). Wenn dem wirklich so ist, dann bedeutet das auch, dass nicht klar ist, ob es überhaupt einen evolutiven Weg gab, auch wenn Ostrom das sicher nicht zum Ausdruck bringen wollte. Padian (2001, 257, 260) nennt die Alternative zwischen Baum- und Cursorialtheorie* (vgl. Junker 2018a) eine „unlösbare Frage“, da die relevanten Vorgänge nicht fossil festgehalten werden könnten (vgl. Padian 2003, 452).2 Beide Theorien seien größtenteils nicht testbar, aber sie seien auch nicht falsifiziert.
 |
| „Unfortunately, we can never know how the different forms of animal flight began.“ |
 |
|
 |
| Abb. 1: Sequenz von hypothetischen Phasen der Evolution des Fluges. I zweibeiniger bodenlebender Archosaurier; II Entwicklung von Anisodaktylie; III Springen auf unteren Ästen von Bäumen und Sträuchern; IV Ansitzen auf Ästen bei vollständiger Ausprägung der Anisodaktylie und Anfangsphase der Reduktion des Schwanzes; V, VI Entwicklung von symmetrischen Federn an den körperfernen Teilen des Schwanzes und der Vordergliedmaßen, zur Unterstützung des Brutverhaltens und des Gleitflugs; VII Bildung von asymmetrischen Federn an den Vorderbeinen und Verkürzung des Schwanzes; VIII Entstehung des aktiven Fluges. (Aus Kurochkin & Bogdanovich 2008, Zeichnung: O. Orekhova-Sokolova, Abdruck mit freundlicher Genehmigung) |
|
Was aber heißt hier überhaupt „testbar“? Und wie kann eine Hypothese zur Flugentstehung falsifiziert werden? Testkriterien, die im experimentellen Bereich Standard sind, kann man nicht anwenden, denn es gibt keine Gesetzmäßig-keiten, aus denen die Entstehung des Vogelflugs abgeleitet werden kann, die man überprüfen könnte. Daher sind alle Theorien zur Entstehung des Vogelflugs weit davon entfernt, experimentell testbar zu sein. Das liegt an der grundsätzlichen Schwierigkeit, dass wir es mit einem hypothetischen Vorgang in der Vergangenheit zu tun haben. Singuläre vergangene Ereignisse sind anders zu testen als regelhaft ablaufende Prozesse, die experimentell untersucht werden können.
Im Anschluss an Bock (2000) ist es sinnvoll, zwischen nomologisch-deduktiven Erklärungen (N-DE) und historisch-narrativen Erklärungen (H-NE) zu unterscheiden (s. Glossar). Tatsächlich kann man nur notwendige Randbedingungen an der Empirie* testen, nicht den hypothetischen Vorgang selbst. Beispielsweise argumentiert Paul (2002, 123), dass die Arborealtheorie (Abb. 1) unterstützt würde, wenn Vogelvorläufer gut ans Klettern angepasst waren. Die Ausbildung von symmetrischen distalen (körperfernen) Federn würde die Sturzflug-Hypothese stützen, während asymmetrische distale Federn die Gleitflug- oder Springer-Hypothese favorisieren würden. Hinweise auf eine schwache Flugmuskulatur sprächen für die Gleitflug-Hypothese.3 Offensichtlich wären solche Indizien keine Belege für den eigentlichen hypothetischen Evolutionsvorgang des Flugerwerbs, denn sie sind nicht spezifisch für diese Hypothese.
Ein zweites Beispiel: Shipman (1998, 189ff.) nennt als Testmöglichkeit dafür, dass Vogelvorfahren Baumbewohner waren, die Frage, ob es in ihrem Lebensraum Bäume gab und ob die Anatomie der betreffenden Arten ein Baumleben ermöglicht. (Damit verknüpft ist die oft schwierige Frage, ob die Einbettungssituation dem damaligen Lebensraum entspricht.) Auch das sind nur Tests auf Randbedingungen hypothetischer Szenarien – mehr nicht. In diesem Sinne äußern sich Dececchi & Larson (2011, 1), dass zwar keine dieser Theorien direkt getestet werden könne, aber dass es möglich sei, eines der Erfordernisse für die Arborealtheorie zu überprüfen, nämlich ob es baumlebende Paraves-Gattungen gebe.4 Die lange Liste der Probleme der jeweiligen Modelle (vgl. Junker 2018a) hat aber gezeigt, dass selbst die Erfordernisse nicht als erfüllt gelten können. Und darüber hinaus sind auch die erforderlichen Randbedingungen oft kaum nachweisbar. Man denke beispielsweise an die Unsicherheiten über die Lebensweise von Archaeopteryx (Leben am Boden oder auf den Bäumen oder beides?5) oder auch an Unsicherheiten bezüglich der damaligen Umwelt.
Welche Tests müsste eine Evolutionshypothese als H-NE „bestehen“? Und wie könnte sie ggf. falsifiziert werden? In der Diskussion werden vor allem Stimmigkeit mit Fossilfunden, Stimmigkeit mit der Phylogenie sowie Kleinschrittigkeit und Selektierbarkeit genannt. Doch diese Kriterien sind entweder nicht als Tests geeignet oder die vorgelegten Modelle bestehen den Test nicht. Das soll im Folgenden gezeigt werden.
Stimmigkeit mit Fossilfunden
Mit diesem Kriterium ist gemeint, dass Fossilformen gefunden werden sollten, die zu den jeweiligen Theorien passen (Martyniuk 2012, 28). So verweisen Befürworter der Baumtheorie auf neuerdings entdeckte baumlebende Theropoden. Umgekehrt wurde früher deren Fehlen oft als Argument gegen die Arborealtheorie* ins Feld geführt (z. B. Gauthier & Padian 1985, 1936; Padian 2001, 2647), wobei zudem auf eher passende bodenlebende Vorläuferformen verwiesen wurde (z. B. Padian 2001, 265).8 Lange Zeit war dagegen nach Auffassung der meisten Paläontologen die Cursorialtheorie* aus dem Rennen, weil keine Theropoden bekannt waren, die eine Furkula (Gabelbein) oder deren mutmaßliche Vorläuferstruktur, das Schlüsselbein, besessen haben (was sich mittlerweile geändert hat) (Hall & Vickaryous 2015, 443). Das Kriterium der Stimmigkeit mit Fossilfunden ist als vorläufiges Ausschlusskriterium durchaus brauchbar. Solange beispielsweise keine baumlebenden, jedoch viele bodenlebende Maniraptoren bekannt waren, bedeutete das mindestens einen Schwachpunkt für die Arborealtheorie.
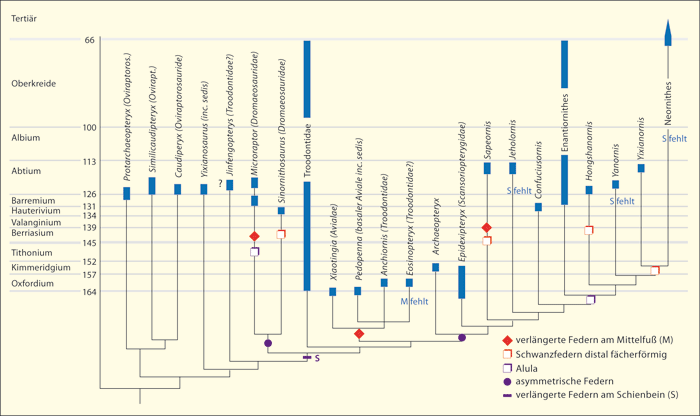 |
| Abb. 2: Gattungen von Dinosauriern mit Konturfedern sowie von Kreide-Vögeln. Das Cladogramm wurde nach Foth et al. (2014) so umgezeichnet, dass die stratigraphische Position der betreffenden Gattungen angegeben werden kann (dicke Balken), und zeigt das Fehlen einer Passung von Phylogenie (cladistischer Verzweigung) und fossiler Abfolge.
Die Gattung Sinornithosaurus ist im Cladogramm zu Unrecht enthalten, da Konturfedern nicht nachgewiesen (und auch nicht wahrscheinlich) sind. Auch bei Epidexipteryx sind keine Konturfedern nachgewiesen; sie werden nur aufgrund indirekter Indizien vermutet; die Gattung besaß allerdings lange, bandartige Schwanzfedern. Die Zuordnung einiger Gattungen zu den Familien ist kontrovers. Es könnten mittlerweile noch weitere Gattungen aufgenommen werden, die das Gesamtbild nicht verändern würden.
|
|
Dieses Kriterium ist dennoch grundsätzlich weich, zum einen, da neue Funde die Situation schlagartig ändern können, aber vor allem, weil es nur einen notwendigen oder wünschenswerten Befund als Stütze für eine Hypothese beschreibt, jedoch nichts über den Vorgang selbst aussagt, der rekonstruiert werden soll. Wenn beispielsweise die Arborealtheorie zum Fossilbefund passen würde, wäre dies kein Beleg dafür, dass es einen evolutionären Prozess von Baumleben zum Flug gegeben hat. Diese Hypothese wäre dann lediglich aufgrund des Fossilbefundes nicht von vornherein ausgeschlossen.
Stimmigkeit mit der Phylogenie
Wenn der Fossilbericht keine Entscheidung über konkurrierende Hypothesen ermöglicht (wie z. B. Padian 2011, 256 meint9), könnte als zweite Möglichkeit geprüft werden, ob die hypothetischen Schritte des Flugerwerbs zu den Cladogrammen (also den stammesgeschichtlichen Rekonstruktionen) passen (Padian 2001, 256; vgl. Abb. 2). Cladogramme werden – ohne Berücksichtigung der stratigraphischen Positionen der berücksichtigten Fossilien – auf der Basis der Verteilung der Merkmale in verschiedenen Taxa konstruiert: An ihnen kann (unter evolutionstheoretischen Voraussetzungen) abgelesen werden, wann welche Merkmale entstanden sein müssten. Dies wird als Testmöglichkeit für die Flugentstehungstheorien genutzt. So wurde in der Vergangenheit als Einwand gegen die Arborealtheorie öfter genannt, dass sie nicht zur Abfolge der für den Flug erforderlichen Merkmale in akzeptierten Phylogenien passe (z. B. Gauthier & Padian 198510; Padian 2001, 255).
Dieses Kriterium ist jedoch ebenfalls vage und noch fragwürdiger als das Fossilien-Kriterium. Denn die Cladogramme ändern sich häufig mit neuen Funden und können daher nicht als hartes Testkriterium gelten. Außerdem sind die Cladogramme gerade im Bereich Theropoden / Urvögel mit Konvergenzen geradezu übersät (Witmer 1991, 460; Peters 2002, 353; Xu et al. 2009, 434; Xu et al. 2010, 3975; Turner et al. 2012, 137)11; wenn aber auch relativ komplexe Merkmale mehrfach unabhängig entstehen können, kann deren Verteilung im Cladogramm keine klare Orientierung in Bezug auf eine Stimmigkeit mit der Phylogenie geben. Damit ist mit diesem Kriterium auch keine schlüssige Bewertung möglich.
Stimmigkeit mit Kenntnissen über Evolutionsmechanismen; kleinschrittige, adaptive Stadien
Plausible H-NE müssen möglichst kleine Schritte in einer hypothetischen Abfolge aufweisen, die durch bekannte evolutionäre Mechanismen (bekannte N-DE) bewältigt werden können (Bock 2000). Die Kritik der einzelnen Modelle hat gezeigt, dass die Kleinschrittigkeit vielfach nicht gegeben ist und dass es bestenfalls unklar ist, wie die bekannten Evolutionsmechanismen die erforderlichen Schritte bewältigen könnten. Man muss sich die Details vor Augen halten, die jeweils erforderlich sind, um einen nächsten selektionspositiven Schritt zu schaffen (vgl. Junker 2016). Das wird kaum gemacht. Bei vielen Szenarien werden zwar einige adaptive Stadien aneinandergereiht, diese sind aber oft durch enorme Sprünge getrennt (vgl. Abb. 3, 4). Es fehlen regelmäßig Detailbetrachtungen darüber, welche Änderungen von Stadium zu Stadium im Einzelnen erforderlich sind, um ein neues adaptives (selektierbares) Stadium erreichen zu können. So aber ist das Kriterium der Kleinschrittigkeit zur Prüfung evolutiver Szenarien viel zu ungenau bzw. die Szenarien erfüllen dieses Kriterium nicht. Beispiele dafür wurden bei der Kritik der einzelnen Modelle ausgeführt (Junker 2018a).
Als weitere Testmöglichkeit wird gelegentlich auch die Existenz oder das Fehlen heutiger Formen mit den in den jeweiligen Modellen postulierten Verhaltensweisen genannt, doch auch dieses Kriterium ist sehr weich (Näheres dazu im Online-Zusatz).
Sind Gegenargumente gegen ein Modell Argumente für seinen Konkurrenten?
In den Diskussionen Pro und Contra Arboreal- und Cursorialtheorie wird für das jeweils favorisierte Modell häufig durch Kritik am Konkurrenten argumentiert. Wenn sich Alternativen gegenseitig ausschließen, ist dieses Verfahren zulässig. Doch in der Diskussion um die Entstehungsweise des Vogelflugs ist das nicht der Fall. Die Tatsache, dass sehr stark mit den Schwächen konkurrierender Modelle argumentiert wird, ist gleichzeitig ein Indiz für die Schwäche des jeweils bevorzugten Modells. Einwände können zudem in der Summe so stark sein, dass alle Modelle extrem unplausibel werden und sich gegenseitig ausschließen. Genau diese Situation scheint bei Flugentstehungshypothesen zuzutreffen und liefert neben anderen Aspekten genügend Anlass, die grundsätzliche Alternative einer Schöpfung in Betracht zu ziehen.
|
Arborealtheorie: Hypothetische Entstehung des Vogelflugs von Bäumen aus. Über Sturzflug und Gleitflug zum Schlagflug (vgl. Abb. 1). Cursorialtheorie: Hypothetische Entstehung des Vogelflugs ausgehend von schnellen Läufern; vom Boden in die Luft. empirisch: auf Erfahrungen (hier: Naturbeobachtung) beruhend. Nomologisch-deduktive Erklärungen (N-DE): N-DE nehmen auf Gesetzmäßigkeiten (gr. nomos = Gesetz) Bezug, aus welchen konkrete testbare Schlussfolgerungen abgeleitet (deduziert) werden. Diese werden dann durch Freilandbeobachtungen oder Laborexperimente überprüft und können ggf. falsifiziert werden. N-DE gelten universell und hängen nicht von der vergangenen Geschichte der Objekte ab, die erklärt werden sollen. Mit N-DE sollen allgemeingütige Aussagen getroffen werden. Historisch-narrative Erklärungen (H-NE): H-NE versuchen das vorhandene Belegmaterial durch ein mutmaßliches historisches Ablaufszenario (eine Erzählung, lat. narratio) zusammenzufügen, wobei solche Szenarien bekannten Gesetzmäßigkeiten nicht widersprechen dürfen. Die Plausibilität von historisch-narrativen Erklärungen hängt also auch davon ab, ob und wie sie nomologisch-deduktive Aspekte (N-DE) berücksichtigen und ob sie auf einer realistischen funktionellen und ökologischen Analyse des Erklärungsgegenstandes fußen. H-NE beschreiben einmalige Ereignisse und beinhalten keine allgemeingütigen Aussagen.
|
|
|
Vogelfedern und Vogelflug als Indiz für Schöpfung
In den Ursprungsfragen der Biologie ist es seit Darwin Standard, grundsätzlich nur Erklärungen zu diskutieren, die dem naturalistischen Paradigma verpflichtet sind. Das heißt: Zugelassen sind nur (rein) natürliche, blinde, nichtgeistige Naturprozesse, die gesetzmäßig oder regelhaft (ggf. durch statistische Gesetzmäßigkeiten) beschreibbar sind, sowie zufällige Randbedingungen. Erklärungen im Rahmen des Naturalismus können lediglich auf drei Faktoren Bezug nehmen: Naturgesetze, (statistisch qualifizierter!) Zufall und plausible Randbedingungen. Jedwede Form einer Schöpfung, d. h. eines zielorientierten Eingreifens, von Zwecksetzungen und von einer überlegten Wahl der eingesetzten Mittel wird nicht berücksichtigt (Widenmeyer & Junker 2016). Die Diskussionen zu den Feder- und Flugentstehungsmodellen haben gezeigt, dass die Erklärungsversuche im Rahmen des Naturalismus bisher faktisch gescheitert sind, auch wenn das von den meisten Wissenschaftlern nicht so gesehen wird, was mit ihrer Festlegung auf den Naturalismus zusammenhängen dürfte. Die Begründungen für das bisherige Scheitern seien hier in Kurzform zusammengefasst:
 |
| Abb. 3: Prums Modell der Federentwicklung. Beachtet man die zahlreichen Details des Federfeinbaus und der Einbettung der Federn im Vogelkörper und ihre Beweglichkeit, wird deutlich, dass 5-6 Stadien viel zu weit auseinander sind, um durch bekannte evolutive Prozesse überwunden werden zu können. Näheres im Text. (1-3 zusammengestellt nach Prum & Brush 2003 und Perrichot et al. 2008; 4-5 von Emily Willoughby, https://emilywilloughby.com) |
|
• Die Federentstehungsmodelle (vergl. Abb. 3) sind sehr vage und nehmen kaum Bezug auf die vielfältigen, detaillierten und miteinander verwobenen Anforderungen für die Flugtauglichkeit der Federn. Modelle, die nur fünf, sechs oder vielleicht acht Abschnitte einer hypothetischen Federevolution beschreiben, sind angesichts der Komplexität des zu erklärenden Gegenstandes völlig unrealistisch und viel zu einfach (Junker 2017a).
• Gegen alle diskutierten Vogelflug-Entstehungsmodelle gibt es eine Fülle von schwerwiegenden Einwänden, von denen die meisten schon für sich alleine das jeweilige Modell in Frage stellen (Junker 2018a).
• Selektionsdrücke, die eine evolutive Entstehung von Federn wahrscheinlich machen, konnten nicht plausibel gemacht werden; entsprechend gibt es keinen Konsens über die hypothetische Erstfunktion von Vogelfedern (Junker 2018b).
• Es sind keine regelhaften oder gar als Naturgesetze beschreibbaren biologischen Zusammenhänge bekannt, die – selbst in kleinen Schritten – eine natürliche Entstehung flugtauglicher Federn, des Vogelflugs und der damit zusammenhängenden Synorganisationen plausibel machen.
Im Rahmen des naturalistischen Ansatzes wird oft argumentiert, es handle sich nur um ein vorläufiges Nichtwissen oder um vorläufig noch offene Fragen bezüglich der Entstehungsvorgänge. Doch woher will man das wissen? Der Wissensfortschritt könnte die Situation genauso gut verschärfen und das ist in der Vergangenheit auch geschehen. So haben z. B. die neueren Forschungen zum Federfeinbau einen äußerst komplizierten Bau der Federn offenbart (Lingham-Soliar & Murugan 2013; Lingham-Soliar 2015; 2017; Laurent et al. 2014; Wang & Meyers 2017). (Zur Problematik der Argumentation mit Nichtwissen und zum Lückenargument siehe Junker 2015.)
|
| Statt des grundsätzlichen Ausschlusses der Erklärungsoption Schöpfung müsste ein Vergleich der verschiedenen Ansätze durchgeführt werden. |
|
|
Vor diesem Hintergrund kann man sagen: Die evolutionären Modelle zur Entstehung von Federn und Flug bleiben nur deshalb im Rennen, weil die grundsätzliche Alternative einer Schöpfung (s. o.) ausgeschlossen wird. Bestünde Offenheit für die Option „Schöpfung“, müsste die Indizienlage im Rahmen eines Vergleichs zwischen Schöpfung und natürlicher Evolution bewertet werden und nicht alleine innerhalb des Naturalismus. Und bei diesem Vergleich hätte der Ansatz „Schöpfung“ folgende Argumente auf seiner Seite:
• Die ausgeprägte Synorganisation der Federstruktur samt Follikel, Federmuskeln, Sinnesorganen in der Haut und der anderen erforderlichen Einrichtungen ist ein klares Schöpfungsindiz. Denn es müssen viele Aspekte und Details über mehrere Ebenen hinweg aufeinander abgestimmt sein, damit Flugtauglichkeit gewährleistet ist.12 Die bekannten natürlichen Mechanismen sind mit der dafür erforderlichen Koordination bei weitem überfordert.
• Die Flugtauglichkeit der Federn – so anspruchsvoll die Voraussetzungen dafür sind – ist aber nur eine notwendige und noch keine hinreichende Voraussetzung für die Flugfähigkeit. Dafür müssen gleichzeitig viele weitere morphologische Strukturen, ausgefeilte Steuerungen und angepasste Verhaltensweisen verwirklicht sein, die wiederum mit den Eigenschaften der Federn abgestimmt sein müssen. Dass dafür enormer planerischer und ingenieurstechnischer Aufwand betrieben werden muss, weiß jeder Flugzeugkonstrukteur.
• Die Flugentstehungshypothesen haben sich nachhaltig als unplausibel erwiesen (Junker 2018a).
• Abgesehen von der Frage nach den Mechanismen kann noch angeführt werden, dass es einen recht klaren Unterschied zwischen Gattungen mit Flaum und Gattungen mit echten (flächigen) flugtauglichen Federn gibt. Zudem erscheint eine Vielfalt verschiedenster Formen ziemlich plötzlich in einem relativ engen geologischen Zeitfenster.
Vom letzten Punkt abgesehen handelt es sich bei der Frage „Schöpfung oder natürlicher Mechanismus“ um sich gegenseitig ausschließende (also echte) Alternativen. Der Schöpfungsansatz geht im Gegensatz zum naturalistischen Ansatz der Evolution davon aus, dass rein natürliche, innerweltliche Prozesse für die Entstehung von Vogelfedern und Vogelflug nicht ausreichen, sondern dass ein geistiger Input wesentlich ist. Für sich gegenseitig ausschließende Sichtweisen gilt: Nachhaltiges Scheitern eines Ansatzes (hier: natürliche Evolution) trotz intensiver Lösungsbemühungen ist zugleich ein Argument für den Alternativansatz (hier: Schöpfung). Das gilt im Falle von Vogelfedern und Vogelflug umso mehr, als hier eine Fülle typischer Design-Indizien nachweisbar ist, die mit dem Fortschritt der Forschung kontinuierlich zugenommen hat. Während die Option „Schöpfung“ also klare Indizien für sich verbuchen kann, steht die Option „Natürliche Mechanismen“ mit ziemlich leeren Händen da. (Zur Definition von „Design-Indizien“ und zur Argumentation damit siehe Widenmeyer & Junker [2016]. Diese grundsätzliche Thematik ist nicht Gegenstand dieses Artikels.)
|
|
|
|
|
Fazit
Die vorausgegangenen Folgen der Artikelserie „Vogelfedern und Vogelflug“ haben gezeigt, dass wesentliche Fragen zur Evolution von Vogelfedern und Vogelflug unbeantwortet sind. Anhand mehrerer Beispiele wurde gezeigt, dass die vorgeschlagenen Evolutionsszenarien zu vage sind und den Details dessen, was zu erklären ist, nicht gerecht werden. Die Entstehung der entscheidenden „evolutionären Neuheiten“ ist auch fossil nicht belegt, auch wenn es viele Mosaikformen mit unterschiedlichsten Merkmalskombinationen gibt. Im Einzelnen kann folgendes Fazit gezogen werden:
 |
| Abb. 4: Komplexes, verschachteltes System von überkreuzten Keratinfasern in der Proteinmatrix im Federschaft und den Federästen in verschiedenen Lagen des Epicortex (seitliche Wände des Schafts). Die Fasern bilden eine ununterbrochene strukturelle Verbindung zwischen dem Cortex des Schafts und der Federstrahlen. (Nach Lingham-Soliar & Murugan 2013; Creative Commons Attribution License) |
|
• Hypothesen zur Entstehung der Vogelfeder müssen deren komplexe Struktur und Synorganisation berücksichtigen. Dabei ist zu berücksichtigen, dass Flugfähigkeit vielfache Wechselwirkung über verschiedene Ebenen hinweg erfordert. Evolutionäre Hypothesen, die diese Synorganisation nicht angemessen beachten, können nicht realistisch sein. So ist die an sich schon sehr komplexe Steuerung der Federbewegungen und ihre Regulation in die Koordination des Fluges eingebettet, wäre zugleich aber wirkungslos, wenn die Federn nicht geeignet strukturiert wären und keinen geeigneten Feinbau besäßen (vgl. Abb. 4). Oder es sei daran erinnert, dass für den Erwerb von Endothermie mehrere anspruchsvolle aufeinander abgestimmte Änderungen erforderlich sind.
• Federn sind gegenüber Reptilschuppen in mehrerer Hinsicht qualitativ andersartige Strukturen und mit diesen bis auf die ersten ontogenetischen Stadien nicht homolog. Ihre hypothetische evolutionäre Entstehung erfordert mehrere Innovationen.
|
| Flugfähigkeit erfordert vielfache Wechselwirkung über verschiedene Ebenen hinweg, die evolutionäre Hypothesen angemessen berücksichtigen müssen. |
|
|
• Das heute weitgehend als Standard akzeptierte Feder-Entstehungsmodell von Prum (1999; Abb. 3) ist mit seinen nur fünf Schritten sehr oberflächlich und übergeht weitgehend die relevanten Fragen, welche Änderungen im Einzelnen von Schritt zu Schritt erforderlich sind und welche Selektionsdrücke diese Schritte begünstigt haben könnten.
• Eindeutige Vorstufen flächiger Federn sind fossil nicht belegt. Zwischen fädigen (und ggf. büscheligen und gering verzweigten) Strukturen und flächigen Federn liegt eine erhebliche morphologische Lücke. Ob man die verschiedenen Formen von Dino-Flaum als Federn bezeichnen kann, ist mindestens fragwürdig und auch eine Frage der Definition. Alle Körperanhänge von Dinosauriern als Federn zu bezeichnen, nur weil aus ihnen evolutiv die Vögel hervorgegangen sein sollen, ist dem Gegenstand nicht angemessen und offenkundig evolutionstheoretisch motiviert und führt zu Zirkelschlüssen. Dino-Flaum jedweder Ausprägung ist weit davon entfernt, einen nennenswerten Beitrag zur Flugfähigkeit zu leisten.
• Gattungen mit flächigen Federn (Konturfedern) sind fossil vor oder etwa gleichzeitig mit Gattungen mit fädigen oder büscheligen Hautanhängen überliefert. Ein Großteil der Gattungen, die Dino-Flaum besitzen und zu Vogelvorläufer-Gruppen gestellt werden, sind stratigraphisch deutlich jünger positioniert als die ältesten Gattungen mit völlig entwickelten, „modernen“ Federn (Archaeopteryx, Anchiornis). Die stratigraphische Abfolge der Gattungen mit Federn oder einfacheren Integumentstrukturen zeigt keine klare Tendenz, die evolutionäre Hypothesen bzw. Abfolgen stützen würde (Junker 2017b). Stratigraphisch erscheint eine erhebliche Vielfalt verschiedenster Formen und Baupläne in einem relativ engen Fenster (vgl. Czerkas & Yuan 2002, 92).13
• Gegen alle Hypothesen zur Entstehung des Vogelflugs – Arborealtheorie, Cursorialtheorie, WAIR-Ansatz (flügelunterstütztes Hochklettern) u. a. – wurden zahlreiche schwerwiegende Einwände formuliert. Selbst wenn einige diese Einwände eventuell entkräftet werden können, sprechen die Fülle und die Schwere der jeweiligen Einwände gegen alle diese Hypothesen. Die einzelnen Hypothesen punkten für sich vor allem durch Einwände gegen ihre(n) Konkurrenten und kaum durch stützende Befunde. Die Kontroverse ist bis heute nicht beendet. Da sie sich ausschließlich im Rahmen des naturalistischen Paradigmas bewegt, kann gefolgert werden, dass sich dieses Paradigma für die Erklärung des Vogelflugs bislang nicht bewährt hat (Junker 2018a).
• Die Selektionsdrücke, die die Entstehung von Vogelfedern begünstigt haben könnten, sind unklar. Es wurde eine Reihe verschiedenster Vorschläge gemacht. Gegen alle Selektionshypothesen können schwerwiegende Einwände geltend gemacht werden (Junker 2018b).
|
Walter Bock, der die hilfreiche Unterscheidung von N-DE (nomologisch-deduktive Erklärungen) und H-NE (historisch-narrative Erklärungen) in die wissenschaftstheoretische Diskussion in der Biologie eingeführt hat, ist der Auffassung, dass ein Schöpfungsansatz aus dem Konzert der konkurrierenden Ansätze herausfalle, weil darin kein Bezug auf N-DE genommen wird. Er schreibt (Bock 2007, 98): „Jede Erklärung historischer Ereignisse, die nicht auf relevanten N-DE basiert, ist nicht wissenschaftlich. Dazu gehören .... Ansätze wie wissenschaftlicher Kreationismus und intelligentes Design, ….“ Damit aber schließt Bock eine mögliche Erklärung grundsätzlich aus, unabhängig von vorliegenden Indizien, und verstößt damit gegen ein Grundprinzip von Wissenschaft, nämlich der Wahrheitsorientierung und Offenheit für verschiedene Antworttypen. Wie gezeigt, liegt überhaupt keine auch nur halbwegs brauchbare H-NE sowohl der Feder- als auch der Vogelflugentstehung vor, u. a. weil die vorgeschlagenen H-NE genau daran scheitern, dass sie nicht auf passenden N-DE basieren (was Bock ja einfordert) bzw. weil empirisch belegte N-DE gegen sie sprechen. Bock (2007, 99) schreibt: „Wenn es keine N-DE gibt oder wenn diese N-DE nicht gut belegt sind, dann liegt die betreffende H-NE außerhalb der Wissenschaft.“ Genau das trifft auf die naturalistisch-evolutionären Feder- und Flugentstehungshypothesen zu. Eine N-DE als Basis der H-NE fehlt. Während evolutionäre Hypothesen jedoch eine oder mehrere N-DE benötigen, beinhaltet der Schöpfungsansatz gerade, dass es eine N-DE nicht gibt, weil ein kreativer Vorgang zugrunde liegt und ein anderer Erklärungstyp benötigt wird. Und da unspezifizierte Schöpfung (Schöpfung als geistig gewirkter Vorgang) und natürliche Evolution sich ausschließende Alternativen sind, bedeutet das Fehlen einer N-DE einen Pluspunkt für den Schöpfungsansatz. Die Forderung einer N-DE als Basis für den Schöpfungsansatz wäre ein logischer Widerspruch und somit eine unsinnige Forderung. Die Forderung nach einem Ausschluss dieses Ansatzes aus der Wissenschaft bedeutet daher nichts anderes als eine Festlegung auf den Naturalismus. Diese Festlegung kann aber durch nichts gerechtfertigt werden.
Wenn aber nur der naturalistische Erklärungsansatz und die damit verbundene naturalistische Weltsicht zugelassen werden, dann ist eine Verifizierung von Hypothesen durch bestätigende Befunde nicht viel wert. Denn wenn es bei H-NE darum geht, im Vergleich zu Konkurrenten die meisten bestätigenden Befunde vorweisen zu können, dann erfordert das eine echte Konkurrenz, nicht ein Rennen nur im Rahmen des einen der beiden grundsätzlichen möglichen Ansätze. Erst dann kann man untersuchen, welcher Konkurrent sozusagen die meisten Punkte holt. In einer Frage: Was ist eine Verifizierung durch bestätigende Befunde wert, wenn es nur eine grundsätzliche Hypothese gibt und nur deren Varianten diskutiert werden?
Wenn es nur einen Konkurrenten gibt, kann man nachvollziehen, dass Dietrich-Bischoff (2014) schreibt, die neuen Befunde zur erstaunlich ausgeklügelten Feinstruktur von Federn könnten möglicherweise dazu beitragen, ein evolutionäres Rätsel, die Entstehung des Vogelflugs, zu lösen, oder dass Lingham-Soliar (2017, 6) meint, es sei klar, dass die außergewöhnliche Mikrostruktur der Wand des Schafts und der Äste über die Millionen Jahre der Vogelevolution perfektioniert worden sei. Zunächst aber zeigen die neuen Befunde, dass das Rätsel größer ist als je zuvor, denn die Federn erweisen sich als komplizierter gebaut als bisher bekannt. Und damit steigen auch die Anforderungen an eine Entstehungshypothese.
|
|
|
Literatur
- Bock WJ (2000)
- Explanations in a historical science. In: Peters DS & Weingarten M (Hg) Organisms, Genes and Evolution. Stuttgart, S. 33–42.
- Bock WJ (2007)
- Explanations in evolutionary theory. J. Zool. Syst. Evol. Res. 45, 89–103.
- Burgers P & Chiappe LM (1999)
- The wing of Archaeopteryx as a primary thrust generator. Nature 399, 60–62.
- Czerkas SA & Yuan C (2002)
- An arboreal maniraptoran from northeast China. In: Czerkas SJ (ed) Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal 1. The Dinosaur Museum, Blanding, USA, pp 63–95.
- Dececchi TA & Larsson HCE (2011)
- Assessing arboreal adaptations of bird antecedents: Testing the ecological setting of the origin of the avian flight stroke. PLoS ONE 6(8): e22292; doi:10.1371/journal.pone.0022292
- Dietrich-Bischoff V (2014)
- Wunderwerk Feder. Die Vogelwarte 52, 59.
- Foth C, Tischlinger H & Rauhut OWM (2014)
- New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers. Nature 511, 79–82.
- Gauthier JA & Padian JK (1985)
- Phylogenetic, functional, and aerodynamic analyses on the origin of birds and their flight. In: In: Hecht MK, Ostrom JH, Viohl G & Wellnhofer P (eds) The beginnings of birds. Eichstätt: Freunde Jura-Museums, pp 185–197.
- Hall BK & Vickaryous MK (2015)
- Merrythoughts of the past and present: revisiting the homology of the furcula. In: Bininda-Emonds ORP, Powell GL, Jamniczky HA, Bauer AM & Theodor J (eds) All Animals are Interesting: A Festschrift in Honour of Anthony P. Russell. BIS Verlag, Oldenburg, pp 439–454.
- Kurochkin EN & Bogdanovich IA (2008)
- On the Origin of Avian Flight: Compromise and System Approaches. Biol. Bull. 35, 1–11.
- Junker R (2015)
- Das Design-Argument in der Biologie – ein Lückenbüßer? http://www.wort-und-wissen.de/artikel/a19/a19.pdf
- Junker R (2016)
- Vogelfedern und Vogelflug. 1. Was Evolutionshypothesen erklären müssten. Stud. Integr. J. 23, 75–82.
- Junker R (2017a)
- Vogelfedern und Vogelflug. 2. Modelle zur Entstehung von Federn. Stud. Integr. J. 24, 4–11.
- Junker R (2017b)
- Vogelfedern und Vogelflug. 3. Belegen Fossilfunde eine Evolution? Stud. Integr. J. 24, 68–77.
- Junker R (2018a)
- Vogelfedern und Vogelflug. 4. Modelle zur Entstehung des Vogelflugs. Stud. Integr. J. 25, 20–27.
- Junker R (2018b)
- Vogelfedern und Vogelflug. 5. Selektion von Feder-Vorstufen – Schlüssel zur Erklärung der Entstehung von Federn? Stud. Integr. J. 25, 76–83.
- Laurent CM, Palmer C, Boardman RP, Dyke G & Cook RB (2014)
- Nanomechanical properties of bird feather rachises: exploring naturally occurring fibre reinforced laminar composites. J. R. Soc. Interface 11: 20140961; doi:10.1098/rsif.2014.0961
- Lingham-Soliar T (2015)
- Feather structure, biomechanics and biomimetics: the incredible lightness of being. J. Ornithol. 155, 323–336.
- Lingham-Soliar T (2017)
- Microstructural tissue-engineering in the rachis and barbs of bird feathers. Sci. Rep. 7:45162; doi:10.1038/srep45162
- Lingham-Soliar T & Murugan N (2013)
- A new helical crossed-fibre structure of ?b-keratin in flight feathers and its biomechanical implications. PLoS ONE 8(6): e65849. doi:10.1371/journal.pone.0065849
- Martyniuk MP (2012)
- A field guide to Mesozoic birds and other winged dinosaurs. Vernon, NJ: Pan Aves.
- Ostrom JH (1986)
- The cursorial origin of avian flight. Mem. Calif. Acad. Sci. 8, 73–82.
- Padian K (2001)
- Stages in the origin of bird flight: Beyond the arboreal-cursorial dichotomy. In: Gauthier J & Gall LF (eds) New Perspectives on the Origin and Early Evolution of Birds. New Haven: Peabody Museum of Natural History, pp 255–272.
- Padian K (2003)
- Four-winged dinosaurs, bird precursors, or neither? BioScience 53, 450–452.
- Paul GS (2002)
- Dinosaurs of the air: The evolution and loss of flight in dinosaurs and birds. Baltimore: John Hopkins University Press.
- Perrichot V, Marion L, Neraudeau D, Vullo R & Tafforeau P (2008)
- The early evolution of feathers: fossil evidence from Cretaceous amber of France. Proc. R. Soc. 275B, 1197–1202.
- Peters DS (2002a)
- Anagenesis of early birds reconsidered. Senckenbergiana lethaea 82, 347–354.
- Prum RO (1999)
- Development and evolutionary origin of feathers. J. Exp. Zool. 285, 291–306.
- Prum RO & Brush AH (2003)
- Zuerst kam die Feder. Spektr. Wiss. 10/03, 32–41.
- Shipman P (1998)
- Taking Wing: Archaeopteryx and the evolution of bird flight. New York: Simon and Schuster.
- Turner AH, Makovicky PJ & Norell MA (2012)
- A review of dromaeosaurid systematics and paravian phylogeny. Bull. Amer. Mus. Nat. Hist. 371, 206 pp.
- Wang B & Meyers MA (2017)
- Light like a feather: A fibrous natural composite with a shape changing from round to square. Adv. Sci. 2017, 4, 1600360.
- Widenmeyer M & Junker R (2016)
- Der Kern des Design-Arguments in der Biologie und warum die Kritiker daran scheitern. Internetpublikation, http://www.wort-und-wissen.de/artikel/a22/a22.pdf
- Witmer L (1991)
- Perspectives on avian origins. In: Schultze HP & Trueb L (eds) Origins of the higher groups of tetrapods. Ithaca: Cornell University Press, pp 427–466.
- Xu X, Zhao Q, Norell M, Sullivan C, Hone D, Erickson G, Wang XL, Han FL & Guo Y (2009)
- A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin. Chin. Sci. Bull. 54, 430–435.
- Xu X, Ma QY, Hu DY (2010)
- Pre-Archaeopteryx coelurosaurian dinosaurs and their implications for understanding avian origins. Chinese Sci. Bull. 55, 3971–3977.
|
|
|  |

