|
Beim Gehen des Menschen spielt eine koordinierte Bewegung von Becken, Lumbalregion* und Brustkorb eine wichtige Rolle. Die Drehung des Beckens in der Transversalebene* ermöglicht dem Menschen, längere Schritte auszuführen. Bei dieser Drehung entsteht ein Drehmoment. Eine gleichzeitige Drehung des Brustkorbes in die entgegengesetzte Richtung führt zu einer Verminderung dieses Drehmoments und damit zu einer Verringerung der aufgewendeten Muskelkraft und Energie beim Gehen. Weiterhin wird der Drehimpuls durch das Schwingen der Arme in entgegengesetzter Bewegungsrichtung der Beine beim Gehen verringert. Es wurde seit langer Zeit vermutet, dass die Fähigkeit zu Rotationsbewegungen des Rumpfes von einem menschenähnlichen langen Rumpf und einer engen Taille abhängig ist.
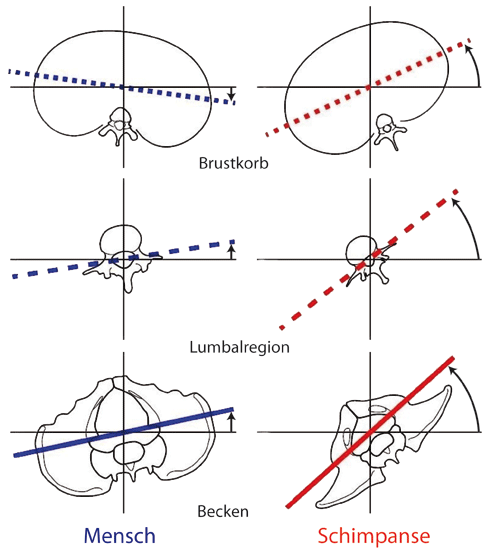 |
| Abb. 1: Rotationsbewegung des Beckens, der Lumbalregion und des Brustkorbes in der Transversalebene von Mensch und Schimpanse bei etwa 50 % des Schrittes. Beim Schimpansen drehen die drei Rumpfsegmente absolut stärker als beim Menschen und in eine Richtung. Beim Menschen rotiert dagegen der Brustkorb in entgegengesetzter Richtung zum Becken und zur Lumbalregion. (Aus Thompson et al. 2015, etwas geändert; CC-BY-Lizenz) |
|
Schimpansen weisen gegenüber dem modernen Menschen eine deutlich verschiedene Rumpfmorphologie auf. Die Beckenschaufeln sind hoch und laden nach außen aus, die Lendenwirbelsäule ist kurz und der untere Brustkorb ist breit. Diese Rumpfmorphologie wurde als starr und unbeweglich angesehen. Diese funktionelle Rigidität impliziert eine sehr begrenzte Bewegungsmöglichkeit zwischen dem Becken und dem Brustkorb in allen anatomischen Ebenen und verhindert eine Rotation des Beckens um die eigene Längsachse des Körpers gegenüber dem Brustkorb in entgegengesetzter Richtung während einer zweibeinigen Fortbewegung.
Thompson et al. (2015) haben diese Annahme geprüft. Sie führten eine dreidimensionale (3D) kinematische Untersuchung der Bewegung des Rumpfes in der transversalen Ebene beim bipeden* Gehen von zwei männlichen Schimpansen sowie von jeweils fünf männlichen und weiblichen Menschen durch. Der Rumpf wurde in drei anatomische Segmente (Brustkorb, Lumbalregion, Becken) unterteilt und die absolute Gesamtbewegung für jedes Segment und die relative Bewegung der Segmente gegeneinander untersucht.
Der Schimpanse dreht alle drei Rumpfsegmente in die gleiche Richtung. Beim Menschen rotieren dagegen Becken und Lumbalregion in eine Richtung und der Brustkorb in die entgegengesetzte Richtung (Abb. 1).
Die Schimpansen zeigen beim bipeden Gehen eine signifikant absolut stärkere Drehbewegung aller drei Segmente im Vergleich zum Menschen. Am stärksten rotiert das Becken, gefolgt von der Lumbalregion und dem Brustkorb.
 |
| Beim zweibeinigen Gehen dreht der Mensch den Brustkorb gegenüber dem Becken in entgegengesetzte Richtung, der Schimpanse dagegen in gleiche Richtung. |
 |
|
Relativ gesehen erfolgt beim Schimpansen die stärkste Rotation des Beckens gegenüber dem Brustkorb, gefolgt von der Lumbalregion gegenüber dem Brustkorb und am geringsten dreht das Becken gegenüber der Lumbalregion.
Bei Mensch und Schimpanse ist das Ausmaß der relativen Bewegung zwischen Becken und Brustkorb und zwischen Becken und Lumbalregion ähnlich, aber zwischen Lumbalregion und Brustkorb ist die Drehung beim Schimpansen signifikant größer als beim Menschen.
Bei Mensch und Schimpanse rotiert der Brustkorb in Beziehung zum Becken maximal kurz nach dem Aufsetzen der Beine.
Zusammenfassend ist festzustellen, dass Menschen und Schimpansen den beim bipeden Gehen auftretenden Drehimpuls, der hohe Muskelaktivitäten und damit Energiekosten verursacht, durch Rotationsbewegungen des Rumpfes verringern. Dabei treten beim Schimpansen gegenüber dem Menschen absolut größere Drehungen aller Rumpfsegmente auf. Beim Menschen rotieren Becken und Lumbalregion gegenüber dem Brustkorb in entgegengesetzter Richtung, während beim Schimpansen alle Rumpfsegmente in gleicher Richtung drehen. In der Studie von Thompson et al. (2015) wurde die Bewegung der Arme bzw. Vorderextremitäten, die beim bipeden Gehen beim Menschen den Drehimpuls zusätzlich vermindern, nicht untersucht.
Dreidimensionale (3D) kinematische Untersuchungen von Ogihara et al. (2010) haben beim Japanmakak (Rotgesichtsmakake) überraschenderweise auch eine Rotation zwischen Becken und Brustkorb mit ähnlichen Messwerten wie beim Schimpansen nachgewiesen. Ogihara et al. (2010) verwendeten allerdings weniger detaillierte Marker als Thompson et al. (2015). Beim Modell von Ogihara et al. (2010) fehlen zwar direkte Marker am Brustkorb, aber die Autoren haben eine Rotation von Becken und Brustkorb in gleicher Richtung festgestellt, wobei die absolute Drehung des Brustkorbs vielleicht geringer ist als beim Schimpansen (Thompson et al. 2015). Diese Befunde zeigen, dass eine Rotation des Beckens gegenüber dem Brustkorb in gleicher Richtung beim bipeden Gehen nicht nur bei Großaffen, sondern auch bei Tieraffen vorkommt.
Thompson et al. (2015) haben nachgewiesen, dass Schimpansen keinen starren Rumpf besitzen, wie dies vorher aus den knöchernen Merkmalen geschlossen wurde. Zu diesen Merkmalen gehören eine kurze Lendenwirbelsäule mit Einschluss des letzten oder der beiden letzten Wirbel zwischen den hohen iliacalen Schaufeln, ein nach lateral ausladender unterer Brustkorb sowie dorsal gelegene Beckenschaufeln.
|
| In der Paläanthropologie sind funktionelle Schlüsse ausgehend von Knochen oft spekulativ. |
|
|
Der Nachweis von ausgeprägten Rotationsbewegungen des Rumpfes bei nichtmenschlichen Primaten während des zweibeinigen Gehens ist sehr lehrreich für die paläanthropologische Forschung, denn man hat vorher aufgrund der Knochenmerkmale auf den gegenteiligen Befund geschlossen. Dieses Beispiel demonstriert erneut (siehe z.?B. Brandt 2015a), wie spekulativ funktionelle Schlüsse sein können, wenn sie allein von Knochenmerkmalen ausgehen. In der Paläanthropologie, wo leider oft nur Knochenüberreste als Untersuchungsmaterial vorliegen, sind solche Schlüsse aber die Regel. Beispielsweise werden von einem einmaligen knöchernen Merkmalsmosaik ganz spezifische Schlüsse bei Australopithecus sediba und Homo naledi zur Fortbewegung und Manipulationsfähigkeit gezogen. Der Autor (Brandt 2014, 2016) hat den hochgradig spekulativen Charakter dieser Schlüsse bei beiden frühen Homininen aufgezeigt. Paläanthropologen sollten aus diesen Gründen bei der Interpretation knöcherner Überreste früher Homininen zurückhaltender sein.
|
biped: zweibeinig. Lumbalregion (Lendenregion): untere rippenfreie Region des Rumpfes. Transversalebene (Axialebene, Horizontalebene): Ebene senkrecht zur Längsachse des Körpers; Drehungen des Körpers erfolgen in dieser Ebene.
|
|
