 |
Vögel – gefiederte Dinos?
von André A. Weller
Studium Integrale Journal
9. Jahrgang / Heft 1 - Mai 2002
Seite 37 - 39
|
| |
 |
| Abb. 1: Compsognathus | |
Seit der Entdeckung des berühmten fossilen „Urvogels“ Archaeopteryx in den 60er Jahren des 19. Jahrhunderts wurde die stammesgeschichtliche Herkunft der Vögel kontrovers diskutiert. Im Mittelpunkt stand dabei häufig die Frage nach der Flugfähigkeit von Archaeopteryx, insbesondere hinsichtlich einer vermuteten Abstammung von zweibeinig laufenden Dinosauriern (Theropoden, z.B. Compsognathus; nach späterer Ansicht Thecodonten; HEILMANN 1926). Basierend auf den frühen anatomisch-morphologischen Studien HUXLEYs im 19. Jahrhundert wurde diese Vorstellung bis in die jüngste Vergangenheit wiederholt von Taxonomen bzw. Paläozoologen aufgegriffen und eine gute Flugleistung zugunsten einer theropoden Herkunft bezweifelt (z.B. BAKKER 1975, OSTROM 1985). Demgegenüber haben andere Evolutionsbiologen bereits frühzeitig auf die „modernen“ Gefiedereigenschaften von Archaeopteryx einschließlich eines gut entwickelten Flugvermögens hingewiesen (vgl. HEINROTH 1926), so daß dessen Rolle als Bindeglied in der frühen Vogelevolution grundsätzlich in Frage gestellt werden muß (FEDUCCIA & TORDOFF 1979). Auch die Anatomie der Füße und Klauen (stärker gebogen als bei Laufvögeln) weist Archaeopteryx als Vertreter der modernen, flugfähigen und baumlebenden Vögel aus (s. z.B. FEDUCCIA 2001a).
Ungeachtet der umstrittenen stammesgeschichtlichen Position von Archaeopteryx haben andere kürzlich entdeckte Fossilien aus der Kreidezeit, die sog. enanthiornithinen Vögel, die Diskussion um den Ursprung der Vögel wiederbelebt. Die neuen Funde zeichnen sich gegenüber – gleichzeitig auftretenden(!) – modernen Formen durch zahlreiche andersartige, spezielle Merkmale aus, beispielsweise einen fleischigen Schwanz, ein verlängertes Pygo-styl (aus miteinander verschmolzenen Wirbeln bestehendes letztes Glied der Schwanzwirbelkette), einen modifizierten Kiel des Brustbeins und vor allem die entgegengesetzte Stellung des Tarsometatarsus (Laufknochen). Einige dieser Gattungen, z.B. Cathyornis, ähneln dem jurassischen Archaeopteryx in wesentlichen Kennzeichen (z.B. in der Schwanzmorphologie). Ein breites Mosaik „primitiver“ und „moderner“ Merkmale charakterisiert die wohl bekanntesten, weil zahlenmäßig am häufigsten gefundenen Vertreter dieser Epoche: die Arten der Gattung Confuciusornis. Einige Paläontologen leiten sie ebenfalls von theropoden, bodenlebenden Sauriern ab (PADIAN & CHIAPPE 1998, OLSON 2000). Demgegenüber hält FEDUCCIA (2001a) eine solche Herkunft (vgl. OSTROM 1973) aus mehreren Gründen für äußerst unwahrscheinlich:
| |
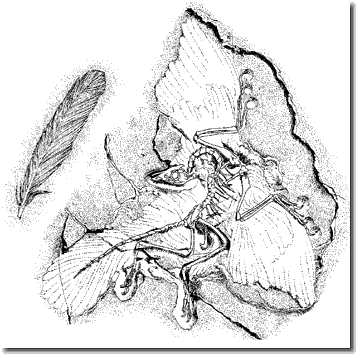 |
| Abb. 2: Archaeopteryx lithographica, der geologisch früheste Vogel, daneben die separat gefundene früheste Feder. (Solnhofener Plattenkalk, Oberjura; Südliche Frankenalb) | |
- Flugfähigkeit ist bei Wirbeltieren an baumbewohnende Vorfahren gekoppelt, außerdem sind Theropoden schon aus aerodynamischen Gründen „ungeeignete“ Vorläufer.
- Zeitlich gesehen treten Dinosaurier, die aufgrund ihres Bau als potentielle Vogel-Vorläufer in Frage kommen, offenbar viel später auf (Altersangabe ca. 120 Ma und jünger) als der bereits relativ moderne Archaeopteryx (ca. 150 Ma). Dazu gehören die den Vögeln – oberflächlich gesehen – am meisten ähnelnden Dromaeosaurier (z.B. Bambiraptor) aus der Kreidezeit, die aber eher als Reliktlinie anzusehen sind.
- Die anatomischen Eigenschaften theropoder Saurier sind kaum auf den Bauplan der Vögel – einschließlich ursprünglicher Formen – übertragbar („character mismatch“), d.h. ein hypothetischer evolutiver „Umbau“ ist aus konstruktiven Gründen nicht möglich. Augenfällige Unterschiede bestehen z.B. in der Konstruktion des Schädels, speziell im Bau der Zähne und ihrer Entwicklung bei den wenigen bezahnten Vogelformen (u.a. sog. Zahnvögel, Hesperornis), die eher denen der Krokodile gleichen. Auch die Vogelhand scheint aufgrund einer unterschiedlichen embryonalen Herkunft der ausgebildeten Handknochen nicht mit derjenigen der Saurier (Finger I-II-III gegenüber II-III-IV bei Vögeln) homologisierbar zu sein, obwohl beide jeweils drei Finger aufweisen – ein zentrales Problem der Vogel-Dino-Abstammungstheorien (vgl. BURKE & FEDUCCIA 1997, FEDUCCIA 2001b). Andere Autoren schließen allerdings eine Verschiebung der Gließmaßenanlagen in der Embryonalentwicklung bei Vögeln nicht aus (u.a. WAGNER & GAUTHIER 1999, GALIS 2001). Ein solcher, sog. homöotischer Austausch würde dazu führen, daß eine strukturelle Anlage (z.B. Finger I) durch eine benachbarte gleichwertige Anlage (Finger II) ersetzt wird und somit quasi eine neue „Identität“ bekommt. Dafür verantwortlich sollen aufeinanderfolgend aktivierte Gene sein, die entweder die Anzahl der Finger (Shh-Gen) bzw. deren Identität (Bmp-Gene) regulieren (DROSSOPOULOU et al. 2000). Die benötigten Mechanismen sind jedoch hypothetisch.
- Die Existenz befiederter Dinosaurier konnte bisher nicht zweifelsfrei nachgewiesen werden. Allerdings gibt es auch hierzu konträre Hypothesen (vgl. ROSSMANN in dieser Ausgabe von SIJ).
- Beim Übergang Kreide-Tertiär starben zahlreiche Vogelformen ebenso wie sämtliche Dinosaurier aus (durch einen Meteoriteneinschlag?). Demgegenüber treten zahlreiche rezente Vogelgruppen plötzlich in Ablagerungen des Paläozän und unteren Eozän auf, vergleichbar der Artenexplosion bei Säugern (FEDUCCIA 1995).
 |
| Anhand des fossilen Belegmaterials ist ein Vorläufer-„Dino“ als Stammvater aller Vögel momentan nicht in Sicht. |
 |
|
Fazit. Zwar wird eine Abstammung der Vögel von Sauriern (baumbewohnender, flug- bzw. gleitfähiger Typus) auch von FEDUCCIA nicht grundsätzlich ausgeschlossen. Allerdings erschweren widersprechende Befunde, z.B. über die Identität morphologischer Strukturen (Vogelhandknochen), eine Interpretation stammesgeschichtlicher Zusammenhänge. Anhand des bekannten fossilen Belegmaterials ist aber ein Vorläufer-„Dino“ als Stammvater aller Vögel momentan nicht in Sicht.
|

Literatur
- BAKKER RT (1975)
- Dinosaur renaissance. Scient. Amer. 232, 58-78.
- BURKE AC & FEDUCCIA A (1997)
- Developmental patterns and the identification of homologies in the avian hand. Science 278, 666-668.
- DROSSOPOULOU G, SANZ-EZQUERRO JJ, NIKBAKHT N, MCMAHON AP, HOFMANN C & TICKLE C (2000)
- A model for anteroposterior patterning of the vertebrate limb based on sequential long- and short-range Shh signalling and Bmp signalling. Development 127, 1337-1348.
- FEDUCCIA A (1995)
- Explosive evolution in Tertiary birds and mammals. Science 267, 637-638.
- FEDUCCIA A (2001a)
- The problem of birds origin and early avian evolution. J. Ornithol. 142, Sonderheft 1, 139-147.
- FEDUCCIA A (2001b)
- Digit homology of birds and dinosaurs: accommodating the cladogram. Trends Ecol. Evol. 16: 285-286.
- FEDUCCIA A & TORDOFF HB (1979)
- Feathers of Archaeopteryx indicate aerodynamic function. Science 203, 1021-1022.
- GALIS F (2001)
- Digit identity and digit number: indirect support for the descent of birds from tetropod dinosaurs. Trends Ecol. Evol. 16: 26.
- HEILMANN G (1926)
- The origin of birds. Witherby, London.
- HEINROTH O (1923)
- Die Flügel von Archaeopteryx. J. Ornithol. 71, 277-283.
- OLSON SL (2000)
- Review of: Chiappe LM et al. (1999) Anatomy and systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of northeastern China. Auk 117, 836-839.
- OSTROM JH (1973)
- The ancestry of birds. Nature 24, 136.
- OSTROM JH (1985)
- The meaning of Archaeopteryx. In: HECHT MK, OSTROM JH, VIOHL G & WELLNHOFER P (eds) The beginning of birds. Eichstätt, pp 161-178.
- PADIAN K & CHIAPPE LM (1998)
- On the origin of birds and their flight. Scient. Amer. 278, 38-47.
- ROSSMANN T (2002)
- Befiederte oder behaarte Dinosaurier? Stud. Int. J. 8, xx-xx.
- WAGNER GP & GAUTHIER JA (1999)
- 1, 2, 3 = 2, 3, 4: a solution to the problem of the homology of the digits in the avian hand. Proc. Natl. Acad. Sci. USA 96, 5111-5116.
|
|  |
