 |
 Studium Integrale Journal
Studium Integrale Journal
25. Jahrgang / Heft 2 - Oktober 2018
Seite 119 - 129
|
| |
|
Eine der vielen faszinierenden Fähigkeiten der Lebewesen ist es, passend auf Umweltreize und Umweltänderungen durch Änderungen in Anatomie, Physiologie oder Verhalten reagieren zu können. Man spricht von Plastizität. Darunter wird die Fähigkeit von Organismen verstanden, auf der Basis desselben Genotyps (Erbguts) mehrere Phänotypen (äußere Form, Physiologie u. a.) als Reaktion auf Umweltreize ausbilden zu können. Beispiele wie die Erhöhung der Anzahl der Roten Blutkörperchen im Blut abhängig vom Sauerstoffgehalt der Luft oder die Bildung einer dickeren Hornhaut bei mechanischer Beanspruchung sind allgemein bekannt.
Plastizität ermöglicht angepasste Reaktionen bei regelmäßig, gelegentlich oder nur ausnahmsweise auftretenden Umweltänderungen. Drei Fähigkeiten sind dafür unverzichtbar: Messung von Umweltreizen, Vergleich mit Sollwerten und passende Reaktionen. Whitman & Agrawal (2009, 20) stellen zum erforderlichen Aufwand für plastische Reaktionen fest: „Zwischen der Reizaufnahme und der Ausprägung des Phänotyps können Dutzende von Schritten liegen, die durch Hunderte von Genen und zahllose Umweltfaktoren und physiologische Faktoren beeinflusst werden“ (Hervorhebung hinzugefügt). Fehlt eine von den drei genannten Voraussetzungen, ist eine plastische Reaktion nicht möglich. Jede dieser drei Fähigkeiten ist anspruchsvoll (z. B. werden „Messinstrumente“ benötigt). Die Fähigkeit zur plastischen Reaktion ist kein gewöhnliches Merkmal wie etwa die Form eines Knochens oder die Anzahl von Blütenblättern, sondern erfordert die Existenz eines Variations- und Anpassungsprogramms sowie geeignete Effektorsysteme, die im Wesentlichen in fertiger Form vorhanden sein müssen. Es ist daher gerechtfertigt, das Vorkommen von Plastizität als Design-Indiz, also als Hinweis auf einen Schöpfer, zu interpretieren (Junker 2014).
 |
| Abb. 1: Das Kriechende Fingerkraut (Potentilla reptans). (CC BY-SA 3.0) |
|
Eine schon länger bekannte plastische Fähigkeit bei Pflanzen sind Strategien zur Vermeidung bzw. Verringerung von Beschattung bzw. passende Reaktionen bei unvermeidbarer Beschattung. Drei Strategien werden beobachtet: Konfrontation, Toleranz und Konkurrenzvermeidung. So erkennen Pflanzen an der Lichtmenge und an den Anteilen verschiedener Wellenlängen des Lichts sowie ggf. durch Berührungsreize und chemische Stoffe die Anwesenheit und Position von konkurrierenden Pflanzen. Reaktion durch Konfrontation erfolgt durch verstärktes Höhenwachstum, um über die Konkurrenten hinauszuwachsen. Eine „tolerante“ Reaktion besteht in der Veränderung der Blattstruktur, z. B. in der Vergrößerung der Blattfläche auf Kosten der Blattdicke, wodurch bei gleichem Materialaufwand das einfallende Licht effektiver genutzt werden kann. Zur dritten Strategie sind Pflanzen in der Lage, die Ausläufer bilden können, wie sie besonders bei der Erdbeere bekannt sind. Dadurch können die Pflanzen einfach seitlich aus dem engeren Umfeld ihrer Konkurrenten herauswachsen und somit die Konkurrenz vermeiden.
Ein Team von Forscherinnen der Universität Tübingen hat nun anhand des Kriechenden Fingerkrauts (Potentilla reptans) untersucht, ob Pflanzen zwischen diesen Reaktionsmöglichkeiten je nach Situation wählen und damit passend auf Größe und Dichte der Konkurrenz reagieren können (Gruntman et al. 2017). Wie der Artname andeutet, sind diese Pflanzen in der Lage, seitlich zu wachsen und Ausläufer zu bilden, sodass ihnen alle drei Reaktionsmöglichkeiten bei veränderter Beschattung zur Verfügung stehen. Die Wissenschaftlerinnen konstruierten verschiedene Umgebungen und variierten diese, um unterschiedlichen Lichteinfall und unterschiedliche vorrangig vorhandene Wellenlängen und auch deren Änderung zu simulieren. Ihre Ergebnisse zeigten, dass die Pflanzen zwischen den drei verschiedenen Strategien hin und her springen können, je nachdem, welche plastische Reaktion gerade aufgrund der Veränderung des Lichteinfalls am passendsten war. Das Kriechende Fingerkraut ist also gleichsam in der Lage, Entscheidungen zu treffen.
In einer Pressemeldung der Universität Tübingen werden die Ergebnisse zusammengefasst: „Es investierte ins Längenwachstum, wenn die Simulation kurz- und dichtwachsende Nachbarspflanzen vorgab – also Konkurrenten, die sich nicht seitwärts umgehen ließen, aber klein genug waren, um in der Höhe überwachsen zu werden. Wurden hoch- wie auch dichtwachsende Nachbarspflanzen simuliert, bei denen weder Flucht noch Konfrontation möglich war, entwickelten die Testpflanzen die höchste Schattentoleranz. Bei hohen, aber licht wachsenden Nachbarpflanzen war die häufigste Reaktion ein seitwärts gerichtetes Wachstum mit Hilfe von Ausläufern“ (https://www.uni-tuebingen.de/uploads/media/17-12-21_Pflanzen__Entscheidungen_dt_02.pdf).
Aus diesen Beobachtungen kann man schließen, dass über die an sich schon bemerkenswerten plastischen Fähigkeiten hinaus die Pflanzen auch einschätzen können, wie dicht und hoch ihre Nachbarn stehen und welche Reaktion daher am besten passt. In Anlehnung an A. Novoplansky sprechen die Autorinnen von „Metaplastizität“, also eine „Plastizität der plastischen Reaktionen“ (Gruntman et al. 2017, 5), und vermuten, dass es eine solche Entscheidungsfähigkeit auch bei anderen Organismen und auch in Bezug auf andere Umweltfaktoren gibt.
Die Fähigkeit, unter verschiedenen Reaktionsmöglichkeiten wählen zu können, ist in einer Umgebung, in der die Konkurrenzverhältnisse sich schnell ändern können, besonders nützlich. Sie kann kaum unterschätzt werden, denn die Pflanzen müssen zunächst über eine genetische, entwicklungsbiologische und biochemische Grundausstattung verfügen, welche die verschiedenen Reaktionen erlauben. Darüber hinaus müssen sie zum einen in der Lage sein, komplexe Informationen über verschiedene Aspekte ihrer Umgebung aufzunehmen und zu verarbeiten, zum anderen müssen sie optimal darauf reagieren und die Reaktionen bei Bedarf immer wieder neu anpassen können. Es ist keine Frage, dass dies einen erheblichen Programmieraufwand erfordert. Die Pflanzen reagieren gleichsam intelligent, als könnten sie verschiedene Einflüsse und ihre Folgen abwägen und somit zukunftsorientiert reagieren: „Sie könnten zu alternativen plastischen Reaktionen übergehen, die entweder Toleranz oder Vermeidung von leichtem Wettbewerb bewirken, und so ihre Reaktion auf die vorherrschenden und zukünftigen Wettbewerbsbedingungen anpassen“ (Gruntman et al. 2017, 2). Da man ihnen keine Denkfähigkeit unterstellen kann, sind diese programmierten und abrufbaren komplexen Fähigkeiten ein klares Design-Indiz.
[Gruntman M, Gross D, Májeková M & Tielbörger K (2017) Decision-making in plants under competition. Nat. Comm. 8:2235, doi:10.1038/s41467-017-02147-2 • Junker R (2014) Plastizität der Lebewesen: Baustein der Makroevolution? Internetartikel, http://www.wort-und-wissen.de/artikel/sp/b-14-2-plastizitaet.pdf • Whitman DW & Agrawal AA (2009) What is Phenotypic Plasticity and Why is it important? In: Whitman DW & Ananthakrishnan TN (eds) Phenotypic Plasticity of Insects: Mechanisms and Consequences. Science Publishers.]
|

|
Einer der Grundmechanismen der Evolution ist Artbildung, also die Aufspaltung einer Art in zwei Tochterarten. Lange Zeit galt im Anschluss an Ernst Mayr als Lehrbuchwissen, dass Artbildung in der Regel eine geographische Trennung (Separation) der Elternart erfordert, damit die getrennten Populationen sich unterschiedlich verändern können, bis sie sich bei späterer Überlappung nicht mehr kreuzen und damit eine Fortpflanzungsisolation erreicht ist. Dieser Prozess soll zudem so langsam verlaufen, dass er auch über mehrere Forschergenerationen nicht beobachtet werden kann (Wagner 2018, 157).
 |
| Abb. 1: Die Hybride (rechts) – Big Birds – aus Geospiza fortis und G. conirostris (links). (Aus Wagner 2018, Abdruck mit freundlicher Genehmigung) |
|
Beide Aspekte sind mittlerweile infrage gestellt, da es starke Indizien dafür gibt, dass Artbildung auch ohne räumliche Trennung und auch in wenigen Generationen möglich ist. Da Artbildung oft auf Spezialisierung beruht, ist es fraglich, ob dieser Prozess auf lange Sicht zu weitergehenden evolutiven Änderungen bis hin zur Entstehung neuer Baupläne führen kann (Junker & Scherer 2017, III.5.1). Vieles spricht dafür, dass Artbildung letztlich in evolutive Sackgassen führt bzw. auf einem Ausschöpfen einer anfangs großen Variationsbreite innerhalb einer Art beruht, was auch sehr gut zum Grundtypmodell der Schöpfungslehre passt.
Besonders gut im Freiland erforscht sind evolutive Prozesse bei den berühmten Darwinfinken der Galapagos-Inseln. Vor einigen Jahren konnte durch Langzeitbeobachtungen eine beginnende Artbildung dokumentiert werden (Grant & Grant 2009; vgl. Junker 2012). Es konnte dabei gezeigt werden, dass die Veränderungen auf dem Ausschöpfen vorhandener Variabilität beruhen.
Nun berichten Lamichhaney et al. (2018) über eine weitere Langzeitstudie bei Darwinfinken, durch die Artbildung durch Hybridisierung nachgewiesen werden konnte (vgl. Wagner 2018). Von 1981 bis 2012 wurde zunächst ein Neuankömmling des Großen Kaktusfinks Geospiza conirostris und in den Jahren danach seine Nachkommen auf der Insel Daphne Major während sechs Generationen beobachtet und deren DNA untersucht. Die Forscher fanden heraus, dass der ungewöhnlich große eingewanderte Vogel von der über 100 km entfernten Insel Española stammen musste. Auf Daphne Major verbandelte er sich mit einem Weibchen des Mittleren Grundfinken (Geospiza fortis). Der daraus entstandene Mischling paarte sich erneut mit G. fortis, und alle weiteren Paarungen erfolgten nur noch unter den neu entstandenen Hybriden (Endogamie) (vgl. Abb. 1).
Der Grund dafür, dass die Hybridenpopulation unter sich blieb, war die ungewöhnliche Körpergröße der Mischlinge, ihr großer und spitzer Schnabel sowie ihr Gesang, der sich deutlich vom Gesang der einheimischen drei Finkenarten unterscheidet. Aufgrund der Fortpflanzungsisolation handelt es sich damit definitionsgemäß um eine neue Biospezies. Die Forscher bemerken, dass die Mischlinge als eigenständige Art betrachtet worden wären, wenn man ihre Vorgeschichte nicht kennen würde.
Die Mischlingspopulation wuchs bis zu acht Brutpaaren und über 30 Individuen an. Trotz Entstehung durch Inzucht weist die neu entstandene Population eine hohe Fitness auf (gemessen an Nachkommenzahl und Überlebensfähigkeit). Aufgrund ihres großen Schnabels sind die Mischlingsvögel in der Lage, die holzigen Früchte der Burzeldorne Tribulus cistoides zu nutzen. Sowohl die Schnabelform als auch der Gesang ermöglichen der neuen Art somit, im Gebiet bereits ansässiger Darwinfinkenarten eine eigene ökologische Nische zu besetzen. Da bei den Darwinfinken der Nachwuchs den Gesang vom Vater lernt, war es entscheidend, dass die Besiedlung von Daphne major durch ein Männchen erfolgte, dessen abweichender Gesang von den Nachkommen unverändert übernommen und erlernt wurde. Dadurch war eine wichtige Voraussetzung für die Abgrenzung von den heimischen Arten erfüllt. Dass aufgrund der besonderen Schnabelform zugleich eine neue ökologische Nische vorhanden war, begünstigte zusätzlich die Entstehung der reproduktiven Isolation und damit die Entstehung einer neuen Art.
Bisher ist der Modus der sog. homoploiden Artbildung (d. h. Artbildung ohne Verdopplung des Chromosomensatzes) kaum beobachtet worden. Wagner (2018) findet deshalb die Studie von Lamichhaney et al. bahnbrechend; sie sei aber auch bedeutsam, weil gezeigt wurde, dass Artbildung sehr schnell innerhalb weniger Generationen erfolgen kann. Wie oft Artbildung durch Hybridisierung vorkommt, kann derzeit nicht abgeschätzt werden; möglicherweise spielt sie eine wichtige Rolle beim Aufbau der Vielfalt innerhalb eines Grundtyps. Es ist gut möglich, dass auch andere der mittlerweile 19 bekannten Darwinfinkenarten aus Hybridisierungen hervorgegangen sind.
[Grant PR & Grant BR (2009) The secondary contact phase of allopatric speciation in Darwin’s finches. Proc. Natl. Acad. Sci. 106, 20141-20148 • Junker R (2012) Schnelle Artbildung live bei Darwinfinken. Stud. Integr. J. 19, 45-47 • Lamichhaney S, Han F, Webster MT, Andersson L & Grant BR (2018) Rapid hybrid speciation in Darwin’s finches. Science 359, 224-228 • Wagner CE (2018) Improbable big birds. Science 359, 157-159]
|
|
Die beiden Wissenschaftler Tobias Lauer und Marcel Weiss vom Max-Planck-Institut für evolutionäre Anthropologie in Leipzig haben neue Daten zum zeitlichen Ablauf der Elster- und Saale-Eiszeit in Mitteleuropa gewonnen. Die Forscher fanden heraus, dass die erste Vergletscherung vor ca. 450000 radiometrischen Jahren (rJ) und damit ca. 100000 rJ früher als bisher angenommen stattgefunden hat. Außerdem zeigten die beiden Wissenschaftler, dass die ersten Menschen Mitteldeutschland bereits vor ca. 400000 rJ und damit früher als bisher allgemein anerkannt besiedelten (Lauer & Weiss 2018). Ein ähnlich hohes Alter könnte aber auch der Fundplatz Bilzingsleben in Thüringen haben (siehe kurze Diskussion bei Lauer & Weiss 2018).
Hinweis auf diese Besiedlung sind zehn einfache Abschlagskerne, 16 Abschläge und drei Werkzeuge, wobei an den Artefakten Kantenschäden und Zeichen der Verrollung nachweisbar sind. Lauer & Weiss (2018) hatten diese kleine Artefaktkollektion im Rahmen von Probeentnahmen für Lumineszenz-Datierungen in den Ablagerungen an der Basis der Saale-Hauptterrasse in Schladebach, Sachsen-Anhalt, entdeckt. Allerdings hatte man aus diesem geologischen Horizont zuvor schon seit der Mitte des 20. Jahrhunderts in Schladebach und Wallendorf mehr als 6.700 Steinartefakte geborgen.
Die neu entdeckten Artefakte von Schladebach sind primär altpaläolithischer Natur, d. h. sie weisen typische Merkmale von Artefakten aus dem Unteren Paläolithikum auf. Nur ein Abschlag weist eine leicht facettierte (bearbeitete) Schlagplattform als Hinweis auf einen (geringen) Trend zu einer mittelpaläolithischen Technologie auf. Außer diesen relativ wenigen Artefakten, die zudem wie schon erwähnt nicht mehr im ursprünglichen Zustand gefunden wurden, gab es keine weiteren Hinweise auf die Existenz der damals lebenden Menschen wie Schlagplätze, Überreste anderer Tätigkeiten des Menschen oder menschliche Knochenüberreste. Lauer & Weiss schreiben nicht, dass man die neu entdeckten Artefakte nur deshalb als Artefakte und nicht als mögliche Geofakte (Naturprodukte) werten kann, weil zuvor schon mehr als 6.700 Artefakte aus diesem geologischen Horizont geborgen wurden. Vielmehr reicht den Archäologen eine kleine Steinkollektion mit Artefaktmerkmalen als Hinweis auf die Existenz von Menschen. Wenn gelegentlich behauptet wird, es würden für den Nachweis menschlicher Tätigkeit zusätzliche Indizien benötigt, so wird dies durch die Forschungspraxis widerlegt.
 |
| Abb. 1: A Schaber aus dem Altpaläolithikum von Schladebach, Sachsen-Anhalt, B Schaber aus dem Pliozän vom Kent-Plateau, England, C Querschaber oder eine gedrungene Spitze aus einem Abschlag aus dem Pliozän von Ipswich, England. (A aus Aus Lauer & Weiss 2018, Supplementary Information, B, C aus Brandt 2011) |
|
Schaut man sich die von Lauer & Weiss (2018) publizierten Artefakte an, so wird der Betrachter an die sogenannten Eolithen erinnert. Eolithen sind Steine aus dem Tertiär mit typischen Artefaktmerkmalen. Sie passen aufgrund ihres hohen Alters nicht in die evolutionären Entstehungsmodelle des Menschen und werden deshalb als Hinweis auf die Existenz von Menschen abgelehnt. Diese Thematik wurde von Brandt (2011) ausführlich thematisiert. In Abb. 1 ist ein von Lauer & Weiss (2018) entdecktes Artefakt zwei Eolithen zur Seite gestellt. Der Leser kann anhand des Vergleichs selbst ein Urteil fällen, ob es sich bei den Eolithen um Steine mit typischen Artefaktmerkmalen handelt.
[Brandt M (2011) Vergessene Archäologie. Steinwerkzeuge fast so alt wie Dinosaurier. Holzgerlingen • Lauer T & Weiss M (2018) Timing of the Saalian and Elsterian glacial cycles and the implications for Middle-Pleistocene hominin presence in central Europe. Sci. Rep. 8:5111. Eine populärwissenschaftliche Zusammenfassung des Originalartikels findet sich unter https://www.mpg.de/11983253/mittelpleistozaen-eiszeiten (Zugriff am 26.3.2018)]
|
|
Bisher galt als Lehrmeinung, dass Schmetterlinge und bedecktsamige Blütenpflanzen „Hand in Hand“ evolvierten. Doch nun zeigen fossile Reste von Schmetterlingsschuppen, dass die einen Saugrüssel tragenden Schmetterlinge sehr viel früher existiert haben. Der Fall zeigt beispielhaft: Stimmige „Geschichten“ sind keine Erklärungen und es muss mit umfangreichen geologisch nicht überlieferten Lebensräumen gerechnet werden.
Schmetterlinge (Lepidoptera) sind der Inbegriff für Symbiose, d. h. eine Vergesellschaftung von Individuen zweier verschiedener Arten, die für beide Partner vorteilhaft ist. Die nektarsaugenden Falter nehmen mit ihrem Saugrüssel beim Blütenbesuch auch Pollen auf bzw. geben ihn ab und ermöglichen auf diese Weise bei vielen Pflanzen die Bestäubung. Sie selber erhalten nahrhaften Nektar aus den Blüten. Was liegt im Rahmen einer evolutionären Weltsicht näher als die Annahme, dass die Entstehung von Schmetterlingen und nektarproduzierenden Blütenpflanzen (Angiospermen) Hand in Hand gingen? Diese Geschichte war bisher Lehrmeinung und mit den bekannten Fossilfunden vereinbar. Allerdings ist der Fossilbericht dieser Insektengruppe sehr dürftig. Der bislang älteste Fund stammte aus der Unterkreide und wird auf 129 Millionen radiometrische Jahre (MrJ) datiert. Molekulare Studien sprechen unter evolutionstheoretischen Voraussetzungen jedoch dafür, dass die Lepidopteren schon sehr viel früher entstanden sind als die Angiospermen, nämlich (je nach verwendeter Methode) im Perm oder in der oberen Trias (> 200 MrJ), während eindeutige Angiospermen-(Makro-)Fossilien erst ab der Unterkreide bekannt sind.
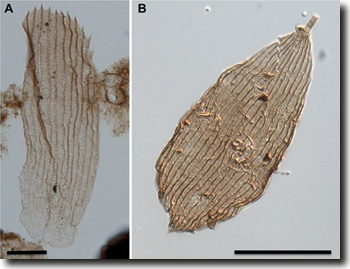 |
| Abb. 1: Präparierte Schmetterlingsschuppen A aus dem Hettangium (Obertrias), B aus dem Rhaetium (Unterjura). Balken: 20 μm. (Aus van Eldijk et al. 2018; CC BY-NC 4.0) |
|
Nun wurden in der Nähe von Braunschweig in einem Sedimentbohrkern aus dem Bereich der Grenze Trias / Jura unter anderen Mikrofossilien zarte, schuppenähnliche Gebilde entdeckt (Van Eldijk et al. 2018). Etwa 70 Schuppen konnten genauer analysiert werden. Die Untersuchung ergab, dass einige von diesen Schuppen Merkmale besaßen, die typisch sind für Flügelschuppen der Gruppe der Glossata, der Motten und Schmetterlinge mit Saugrüssel. Der Besitz solcher Schuppen, die Flügel, Beine und den Körper bedecken, ist das namengebende Merkmal der Lepidoptera – „Schuppenflügler“ (gr. lepis, Schuppe). Besonders die auf den Flügeln sitzenden Schuppen sind taxonomisch informativ, d. h. sie erlauben eine recht genaue Zuordnung zu einer bestimmten Gruppe. Damit ist nun der fossile Nachweis erbracht, dass Schmetterlinge 70 MrJ früher existiert haben als bisher bekannt und ca. 50 MrJ vor dem ersten Nachweis von Angiospermen. Die Geschichte von der Co-Evolution von zwittrigen, nektarführenden Blüten und Insekten mit Saugrüssel passt also nicht mehr zum fossilen Befund.
Wie könnte eine neue „Geschichte“ aussehen? Die Forscher mutmaßen, dass der Saugrüssel anfangs als Trinkwerkzeug evolvierte, mit dem Tropfen von pflanzlichem Saft aufgenommen werden könnten (z. B. Bestäubungstropfen von Nadelbäumen, mit denen Pollen aufgefangen werden, oder bei Verletzungen von Pflanzen austretende Flüssigkeit). Es gibt Hinweise auf große Trockenheit in jener Zeit, weshalb der Besitz eines Saugrüssels vorteilhaft war. Der Nutzen war zuerst einseitig nur beim Insekt. Erst später sei der Saugrüssel dann für die Aufnahme von Nektar mit gegenseitigem Nutzen (Bestäubung) verwendet worden.
Diskussion. Die Umwandlung der zuvor beißenden Mundwerkzeuge in ein Saugorgan wird von Van Eldijk et al. (2018) als evolutionäre Antwort auf Trockenheit beschrieben. Wie die Umwandlung erfolgte, wird nicht thematisiert. Es ist einfach eine neue Geschichte, die an die Stelle der alten getreten ist. Solche Geschichten sind keine Erklärungen, sie können sich jederzeit durch neue Funde als falsch erweisen. Sie sind vage, gehen nicht in Details und wirken geradezu „undarwinistisch“, weil der Eindruck erweckt wird, dass als Folge veränderter Bedürfnisse (hier Trockenheit) passende Reaktionen zielgerichtet möglich seien. Das erinnert sehr an Lamarcks hypothetischen Ansatz, wonach Umweltreize einen direkten Einfluss auf den optimalen und angepassten Gestaltwandel der Organismen ausgeübt haben sollen. Die neue Geschichte ist zudem wenig glaubwürdig, da viele andere Insekten zur gleichen Zeit ohne saugende Werkzeuge auskamen. Warum also sollte sich gerade in der Linie, die die Lepidopteren hervorbrachte, ein Saugrüssel bilden? Das ist völlig unklar, zumal Angebote von Flüssigkeit zum Saugen auch schon vorher vorhanden waren. Sowohl die bisherige als auch die neue Geschichte sind typische Beispiele von „adaptivem Storytelling“. Die bisherige Geschichte ist im Übrigen gar nicht widerlegt, wie behauptet wird. Würde man nämlich in Schichten der Trias Angiospermen finden, würde sie wieder passen. Dafür gibt es sogar Hinweise (Hochuli & Feist-Burkhardt 2013; vgl. Kutzelnigg 2014), auf die die Autoren selber hinweisen (van Eldijk et al. 2018, 5).
In jedem Fall zeigen die neuen Funde, dass mit umfangreichen fossil nicht überlieferten Lebensräumen gerechnet werden muss, da es eine Lücke von 70 MrJ zwischen den stratigraphisch ältesten und zweitältesten fossilen Nachweisen der Lepidopteren gibt. Auch der große zeitliche Versatz zwischen dem erstmaligen fossilen Auftreten von angiospermenartigem Pollen und dazu passenden Makrofossilien deutet auf fossil nicht überlieferte Lebensräume hin.
Die Daten laden darüber hinaus auch dazu ein, über einen prinzipiell anderen Deutungszugang bezüglich der geologischen Überlieferung nachzudenken, der u. a. ohne Evolution und lange Zeiträume als paradigmatischem Rahmen auskommt. Der Autor ist sich bewusst, dass dazu im Rahmen der Schöpfungsforschung bisher nur sehr wenig vorgelegt werden konnte.
[Hochuli PA & Feist-Burkhardt S (2004) A boreal early cradle of Angiosperms. Angiosperm-like pollen from the Middle Triassic of the Barents Sea (Norway). J. Micropalaeontol. 23, 97-104 • Kutzelnigg H (2014) Sind Blütenpflanzen 100 Millionen Jahre älter als bisher angenommen? Stud. Integr. J. 21, 41-43 • Van Eldijk TJB, Wappler T et al. (2018) A Triassic-Jurassic window into the evolution of Lepidoptera. Sci. Adv. 2018;4: e1701568]
|
 |
| Abb. 1: Das Londoner Exemplar von Archaeopteryx mit gut erhaltenen Federn. (Bild: Muséum national d'Histoire naturelle, Paris, Nachbildung, gemeinfrei) |
|
Es dürfte kaum eine fossile Art geben, über die so viel und so kontrovers publiziert wurde wie über Archaeopteryx, den berühmten „Urvogel“. Bis heute gehen die Meinungen über seine Flugfähigkeit auseinander, insbesondere bezüglich der Frage, ob Archaeopteryx aktiv fliegen konnte. Einerseits spricht der Besitz modern anmutender asymmetrischer Federn dafür, dass Archaeopteryx ein aktiver Flieger war, andererseits scheinen einige anatomische Gegebenheiten nicht dazu zu passen, zum Beispiel das Fehlen eines Brustbeinkiels oder die Anatomie des Schultergelenks.
Seit kurzem kann mithilfe einer speziellen Form der Röntgen-Mikrotomographie die Struktur fossiler Knochen auf eine neue Weise untersucht werden, die bisher unzugängliche Einsichten in die histologischen Strukturen ermöglichen. Dennis Voeten und sein Team vom European Synchrotron in Grenoble (ESRF) untersuchten mit dieser Methode drei Archaeopteryx-Fossilien und verglichen die virtuellen Querschnitte der Knochen mit der Knochenstruktur anderer fossiler Arten (auch von Flugsauriern) und heute lebender Formen. Es stellte sich heraus, dass ein zuverlässiger Zusammenhang zwischen der Knochenarchitektur und dem Flugstil hergestellt werden kann. Dies wiederum konnte genutzt werden, um die Flugfähigkeit von Archaeopteryx einzuschätzen. Dessen Flügelknochen, insbesondere der mittlere Teil der Armknochen, zeigen große Gemeinsamkeiten mit den betreffenden Strukturen bei modernen Vögeln und sind deutlich verschieden von den Knochen landlebender Dinosaurier; so ist die Knochenwand bei Archaeopteryx viel dünner als bei letzteren. Besonders gut vergleichbar sind die Knochenstrukturen mit der Situation bei Fasanen. Die Forscher schließen daraus, dass Archaeopteryx ähnlich wie heutige Fasanen gelegentlich aktiv hochflattern konnte, um Hürden zu überwinden oder Feinden auszuweichen. Dagegen waren sie vermutlich keine ausdauernden Flieger.
Andererseits gibt es markante anatomische Unterschiede zu heutigen Vögeln. Hier bleiben für das Verständnis der Art des Fluges bei Archaeopteryx Fragen für die Wissenschaftler offen. Sie vermuten, dass Archaeopteryx auf eine bislang nicht geklärte Weise mit anderer Haltung und anderer Art des Flügelschlags unterwegs war als heutige Vögel.
Die neuen Befunde reihen sich in die Liste neuerer Untersuchungen ein, die gezeigt haben, dass die geologisch (bzw. genauer stratigraphisch) ältesten Vögel bessere Flieger waren als bisher angenommen (vgl. Literatur am Ende). Je mehr Details aufgeklärt werden können, desto eher scheinen Anzeichen von Primitivität zu verschwinden. Das gilt für die sogenannten Gegenvögel (Navalón et al. 2015), für den „Konfuziusvogel“ (Confuciusornis) (Falk et al. 2016) oder auch für die Gattung Anchiornis (Wang et al. 2017), die möglicherweise zu Unrecht zu Dinosauriern statt zu den Vögeln gestellt wird (Junker 2017). Die Aufklärung der mutmaßlich evolutionären Entstehung des Vogelflugs rückt damit in größere Ferne als bisher. Primitive Flugvorstufen sind bisher nicht sicher nachweisbar.
[Falk AR, Kaye TG, Zhou Z & Burnham DA (2016) Laser fluorescence illuminates the soft tissue and life habits of the Early Cretaceous bird Confuciusornis. PLoS ONE 11(12): e0167284. doi:10.1371/journal.pone.0167284 • Junker R (2017) Anchiornis – nur „fast ein Vogel“? Stud. Integr. J. 24, 102-104 • Navalón G, Marugán-Lobón J, Chiappe LM, Sanz JL & Buscalioni A (2015) Soft-tissue and dermal arrangement in the wing of an Early Cretaceous bird: Implications for the evolution of avian flight. Sci. Rep. 5:14864 • Voeten DFAE, Cubo J, de Margerie E, Röper M, Beyrand V, Bure S, Tafforeau P & Sanchez S (2018) Wing bone geometry reveals active flight in Archaeopteryx. Nat. Comm. 9:923, doi: 10.1038/s41467-018-03296-8 • Wang X, Pittman M, Zheng X, Kaye TG, Falk AR, Hartman SA & Xu X (2017) Basal paravian functional anatomy illuminated by high-detail body outline. Nat. Comm. 8:14576, doi: 10.1038/ncomms14576]
|
|
In den Medien erregen Meldungen mit Hinweisen auf Leben irgendwo im All große Aufmerksamkeit. Die Lektüre offenbart dann, dass die Hinweise auf sehr optimistischen Interpretationen von wenigen Befunden beruhen und somit eher Wunschträumen oder Erwartungshaltungen der Öffentlichkeit entspringen. Hier einige aktuelle Forschungsergebnisse und deren Einordnung und Bewertung.
Gebiete in der Atacama-Wüste im Westen Südamerikas (im Süden Perus und in Chile) zählen zu den trockensten Lebensräumen der Erde. Nach den äußerst seltenen Niederschlägen zeigt sich, dass sich auch hier Leben entfalten kann, wenn die entsprechenden Wüstenregionen für begrenzte Zeit aufblühen. Die meiste Zeit aber stellt diese Region enorme Herausforderungen an jegliche Art von Lebewesen.
 |
| Abb. 1: Enceladus, ein großer Saturnmond, ist ein Eismond. Die Aufnahme lässt die vulkanische Aktivität erkennen. Die Fontänen bestehen aus Wassereis. Hinweise auf flüssiges Wasser nähren Spekulationen über günstige Bedingungen zur Lebensentstehung. (Bild: NASA) |
|
Ein umfangreiches internationales Wissenschaftlerteam hat nun untersucht, ob unter den in der Atacama-Wüste vorherrschenden Bedingungen wie extreme Trockenheit, sehr salzige Böden und ausgeprägter Nährstoffmangel mikrobielles Leben dauerhaft möglich ist. Die Forscher sammelten Proben im Jahre 2015 – nach dem letzten nennenswerten Niederschlagsereignis – sowie in den beiden darauffolgenden Jahren (2016 und 2017), in denen keine Niederschläge verzeichnet worden waren. Sie konnten anhand von Proben an verschiedenen geographischen Regionen zeigen, dass sowohl die Biomasse als auch die Vielfalt der Mikroorganismen mit zunehmender Trockenheit abnimmt. Unter den extremen Lebensbedingungen ist es schwierig, Leben überhaupt zuverlässig nachzuweisen. Daher verwendeten die Autoren unterschiedliche Kriterien für den Nachweis von mikrobiellem Leben. Sie konnten vier verschiedene Hinweise auf dauerhaft lebende Mikroben (neben den sehr robusten Sporen, die manche Bakterien bilden) aufzeigen: (a) die physikalisch-chemische Beschaffenheit des Bodens, der durch Pflanzen nach einem der sehr seltenen Regenereignisse nutzbar ist; (b) den Nachweis von Biomolekülen, die auf aktive Zellen hinweisen, wie ATP, Phospholipide, Stoffwechselprodukte und enzymatische Aktivität; (c) messbare Verdopplungsraten von Genomen in Bodenproben und (d) spezifische Zusammensetzung der Mikrobengemeinschaften, abhängig von der Qualität des Bodens und der Tiefe, aus der die Proben genommen wurden.
Aus ihren Befunden leiten die Autoren ein Arbeitsmodell für den Mars ab, obwohl sie davon ausgehen, dass die dort herrschenden Bedingungen noch extremer sind. Sie gehen davon aus, dass, wenn sich je Leben auf dem Mars entwickelt haben sollte, Überreste davon in Nischen unter der Oberfläche zu finden sein könnten. Für die Erde wissen wir, dass es hier Lebewesen gibt, die uns immer wieder mit erstaunlichen Eigenschaften überraschen. Belege dafür, dass es auf dem Mars jemals Leben gegeben hat, fehlen allerdings bisher!
Über die Möglichkeiten außerirdischen Lebens wird nicht nur im Zusammenhang mit dem vergleichsweise erdnahen Planeten Mars spekuliert, auch andere Himmelkörper werden unter astrobiologischen Fragestellungen ins Blickfeld genommen. In einer NASA-Mission (2004-2017) wurden mit der Raumsonde Cassini (und Huygens) Saturn und seine Monde untersucht. Im Rahmen dieser Mission wurden am Südpol des Saturnmondes Enceladus Wasserdampf-Fahnen dokumentiert. Diese Befunde führten bereits 2008 zu Spekulationen über Leben auf diesem Mond (McKay et al. 2008). Weitere Daten wurden als Hinweise auf hydrothermale Prozesse interpretiert, die unter der Eisdecke dieses Himmelskörpers ablaufen könnten. In jüngeren Publikationen werden neben Wasser auch Methan (CH4), Kohlenstoffdioxid (CO2), Ammoniak (NH3), molekularer Stickstoff (N2) und Wasserstoff (H2) nachgewiesen (Waite et al. 2017) und damit die Vermutung hydrothermaler Vorgänge auf Enceladus erhärtet.
Taubner et al. (2018) haben jetzt unter der Annahme, dass zwischen dem als steinern angenommenen Kern und der vereisten Oberfläche des Saturnmondes in flüssigem Wasser geochemische Prozesse bei gemäßigten Temperaturen (< 100 °C) ablaufen, die Idee geprüft, ob ein Teil des Methans biologischen Ursprungs sein könnte. Sie konnten im Labor unter simulierten Enceladus-Bedingungen zeigen, dass Methan produzierende Archaebakterien unter den dort angenommenen Drücken CO2 in CH4 umwandeln. Damit könnte, so schlussfolgern die Autoren, ein Teil des nachgewiesenen CH4 biologischen Ursprungs sein und damit auf Leben hinweisen.
Auch dieser indirekte Hinweis, dass es auf Enceladus möglicherweise Lebewesen geben könnte, ist stark von der Überzeugung geprägt, dass biologische Systeme auch an anderen Orten im Universum unter geeigneten Bedingungen zu erwarten sind, ohne Absicht und entsprechende Steuerung. Für diese Überzeugung gibt es jedoch keinen Anlass und bisher liegt auch kein empirischer Beleg für extraterrestrisches Leben vor.
[McKay CP, Porco CC, Altheide T, Davis WL & Kral TA (2008) The possible origin and persistence of life on Enceladus and detection of biomarkers in the plume. Astrobiology 8, 909-919 • Schulze-Makuch et al. (2018) Transitory microbial habitat in the hyperarid Atacama desert. Proc. Nat. Acad. Sci. USA; doi: 10.1073/pnas.1714341115 • Taubner R-S, Pappenreiter P, Zwicker J, Smrzka D, Pruckner C, Kolar P, Bernacchi S, Seifert AH, Krajete A, Bach W, Peckmann J, Paulik C, Firneis M, Schleper C & Rittmann SK-MR (2018) Biological methane production under putative Enceladus-like conditions. Nat. Comm.9, 748; doi: 10.1038/s41467-018-02876-y • Waite JH et al. (2017) Cassini finds molecular hydrogen in the Enceladus plume: evidence for hydrothermal processes. Science 356, 155-159.]
|
|
Das Phänomen Leben ist auf allen erforschten Ebenen überaus komplex. Selbst die kleinste Einheit des Lebens, die Zelle, besteht aus einer Vielzahl an eng verwobenen und aufeinander angewiesenen Organellen. Als Organellen (kleine Organe) werden zelluläre Untereinheiten bezeichnet, die verschiedene Aufgaben im Haushalt der Zelle übernehmen. In tierischen Zellen sind z. B. Mitochondrien für die Energieversorgung zuständig, Peroxisomen für den Abbau giftiger Nebenprodukte des Sauerstoffs und Lysosomen für den Abbau einer Vielzahl an natürlichen Stoffgruppen, die entweder von außen in die Zelle gelangen oder innerhalb der Zelle erzeugt werden (Nukleinsäuren, Proteine, Fette etc.). In pflanzlichen Zellen sind Chloroplasten für die Energiespeicherung durch Photosynthese unverzichtbar. Alle bisher genannten Organellen haben ungeachtet ihrer hochgradig unterschiedlichen Funktionen eines gemeinsam: Sie weisen alle nach außen eine Abgrenzung auf – eine Membran. Es sind zwar auch einige Organellen ohne Membran bekannt, wie die als „Proteinfabriken“ bezeichneten Ribosomen, doch weisen auch sie eine wohldefinierte, feste Form auf. Eine weitere Gemeinsamkeit aller bisher bekannten Organellen ist die relativ komplexe Zusammensetzung aus vielen verschiedenen Molekültypen.
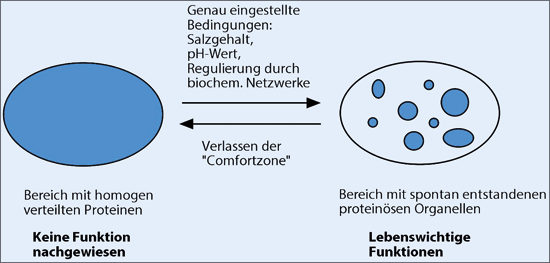 |
| Abb. 1: Schematische Darstellung der Bildung membranloser Organellen. |
|
Anders sieht es dagegen bei einem vor wenigen Jahren entdeckten Typ von Organellen aus, den proteinösen membranlosen Organellen („proteinaceous membrane-less organelles“, PMLO, Uversky 2017). Im Unterschied zu den meisten bisher bekannten zellulären Funktionseinheiten zeichnen sich die PMLO nicht nur durch das Fehlen einer Membran aus, sondern sie sind häufig flüssig. Sie werden als kleine Tröpfchen beschrieben, die sich beim Auftreffen auf Hindernisse stark und umkehrbar verformen können. PMLOs können darüber hinaus Wasser aufnehmen und miteinander fusionieren. Wie es der Begriff „proteinös“ schon andeutet, bestehen die PMLOs hauptsächlich aus Proteinen, wobei sie in einigen Fällen auch Nukleinsäuren enthalten. Von besonderem Interesse ist hierbei die Art der Proteine, die für PMLOs konstitutiv sind: Es handelt sich ausschließlich um intrinsisch unstrukturierte Proteine (IUP); das sind Proteine ohne feste 3D-Struktur, die sich aber häufig durch Multifunktionalität auszeichnen (Schmidtgall 2018). Neben der auffällig selektiven Zusammensetzung der PMLOs sind auch andere Eigenschaften erstaunlich. PMLOs stellen eine bisher ungeahnte Verdichtung biologischer Makromoleküle dar. Im Vergleich zum ohnehin sehr dichten zellulären Milieu (je nach Kompartiment 80-400 g/l Gesamtkonzentration) beträgt die gesamte Konzentration der Makromoleküle innerhalb der PMLOs das 10-300-fache.
Die reversible Entstehung dieser äußerst dichten Tröpfchen geschieht allerdings nur unter ganz präzise eingestellten Bedingungen und einer exakten Kontrolle durch Phasentrennung – vergleichbar der Trennung von Öl und Wasser. Diesen Vorgang beschreibt Uversky in einem kürzlich erschienenen Übersichtsartikel mit folgenden Worten: „Typischerweise ist ein solcher Vorgang komplett umkehrbar, hochgradig kontrolliert und in hohem Maße von den Bedingungen abhängig, d. h. dass die Phasentrennung nicht geschieht, solange die spezifischen Bedingungen dafür nicht eingestellt sind. Hingegen führt ein Verlassen der Komfortzone zu einem kompletten Verschwinden der kondensierten Phase. […] ein ‚jetzt siehst du mich, jetzt nicht mehr’-Phänomen.“
Zu den Faktoren, von denen die beschriebenen Phasenübergänge abhängig sind, zählen nicht nur physikalische Parameter wie der pH-Wert, die Salz- und Makromolekülkonzentration und die Temperatur, sondern auch biochemische Eigenschaften der beteiligten Proteine wie Aminosäuresequenz und die Anzahl an Phosphorylgruppen (PO32-), die erst nach der Translation (Übersetzung der mRNA in Proteine) eingefügt werden.
Ungeachtet ihrer hohen Empfindlichkeit sind PLMO zahlreich und gelten als überaus wichtige Organellen, die in vielen verschiedenen Formen vorkommen und spezifische Funktionen ausüben, die in hohem Maße mit den jeweiligen Prozessen korrelieren. Zu den wenigen bisher bekannten Funktionen gehören die Stressantwort, der Transport von RNA-Molekülen innerhalb des Zellkerns, das Spleißen von mRNA-Molekülen, die Regulierung der Genexpression und die Reaktion auf Schäden an der DNA.
Zusammenfassend kann also konstatiert werden, dass ein neuer Typ von zellulärer Funktionseinheit entdeckt worden ist, der trotz seiner außergewöhnlichen Spezifität und Empfindlichkeit gegenüber einer Vielzahl an Parametern sehr wichtige Funktionen ausübt. Im Hinblick auf evolutionstheoretische Modelle stellt sich die Frage, wie solche empfindlichen Zellbestandteile auf Mutationen reagieren und wie ein derart fein abgestimmtes System durch ein zukunftsblindes Zusammenspiel von Zufall und Notwendigkeit („trial and error“) zustande gekommen sein kann. Bisher deuten jedenfalls einige Befunde darauf hin, dass Mutationen, die die konstitutiven Proteine von PMLO betreffen, zum Verklumpen der dichten, flüssigen Kügelchen führen können. Dies wäre nicht überraschend, da IUP schon öfter im Zusammenhang mit schweren Erkrankungen gebracht worden sind, die aus Protein-Verklumpungen resultieren (z. B. neurodegenerative Erkrankungen). Auch der Umstand, dass die PMLO einer intensiven Regulierung durch die biochemischen Netzwerke der Zelle bedürfen, spricht gegen die Möglichkeit einer schrittweisen, ungerichteten Entwicklung der rätselhaften proteinösen Organellen. Es erstaunt jedenfalls nicht, wenn Uversky (2017) im Hinblick auf die PMLO von „einer Magie der biologischen flüssig-flüssig-Phasenübergänge“ spricht.
[Schmidtgall B (2018) Intrinsisch unstrukturierte Proteine. Stud. Integr. J. 25, 84-91 • Uversky VN (2017) Intrinsically disordered proteins in overcrowded milieu: membrane-less organelles, phase-separation and intrinsic disorder. Curr. Opin. Struct. Biol. 44, 18-30]
|
 |
| Abb. 1: Mausohr (Myotis myotis) im Quartier in einer Brücke. (Foto: MissMhisi,CC BY-SA 4.0) |
|
Eines der rätselhaften Phänomene des Lebens ist das Altern. Die Hoffnung ist, dass durch ein Verständnis der Alterungsprozesse auch Einfluss auf deren Geschwindigkeit genommen werden könnte. Möglichst lange jung zu bleiben, ist ein uralter Traum der Menschheit. Bibelleser kennen die hohen „biblischen“ Alter der Urväter mit dem Altersrekord von 969 Jahren des auch sprichwörtlich genutzten Methusalem. Könnten solche hohen Alter realistisch sein oder sind sie eine biologische Unmöglichkeit?
Von überraschender Seite wurden zu dieser Frage kürzlich hochinteressante Befunde publiziert. Eine Forschergruppe (Foley et al. 2018) berichtete über Untersuchungen des Alterns bei verschiedenen Arten von Fledermäusen und fand dabei heraus, dass der Vorgang des Alterns beim Großen Mausohr (Myotis myotis, Abb. 1) sehr viel langsamer als bei anderen Fledermausarten verläuft. Bekanntlich gibt es bei Säugetieren einen ungefähren Zusammenhang zwischen Körpergewicht und Lebenserwartung. Je geringer das Körpergewicht, desto geringer das Lebensalter. So haben Mäuse eine Lebenserwartung von wenigen Jahren, Hunde werden bis zu ca. 15 Jahre alt, große Säugetiere ca. 50 Jahre. Der Mensch fällt somit deutlich aus dem Rahmen. Aber im Vergleich zum Körpergewicht ist das noch nichts gegen das Große Mausohr. Obwohl es nur wenige Gramm schwer wird und entsprechend nur wenige Lebensjahre zu erwarten hätte, erreichen die Tiere oft ein Lebensalter von 35 Jahren, ungefähr zehnmal so viel, wie aufgrund ihres Gewichts zu erwarten wäre.
Was ermöglicht diesen enormen Ausreißer? Schon lange wird ein Zusammenhang zwischen dem Altern und der Länge der Endstücke der Chromosomen, der Telomere, vermutet. Bei jeder Zellteilung werden die Telomere verkürzt, bis schließlich keine Zellteilungen mehr möglich sind. Allerdings verzögert die Tätigkeit des Enzyms Telomerase die Verkürzung, indem es die Telomere in beschränktem Maße wiederherstellen kann, was jedoch mit zunehmendem Alter nachlässt. Entsprechend können Mutationen der Telomerase zu vorzeitigem Altern führen. Beim berühmten Klon-Schaf Dolly waren die Telomere schon bei der Geburt verkürzt. Die geringe Lebensspanne und der früh einsetzende Altersprozess von Dolly dürften damit zusammenhängen und dieser Befund wird auch als Stütze für die Telomer-Hypothese des Alterns angesehen.
Die Verhältnisse bei den Fledermäusen erwiesen sich allerdings als komplizierter. Zwar konnte ein Zusammenhang mit dem Ausbleiben der Telomerverkürzung bestätigt werden, doch die Ursachen dafür sind vielfältig und bestehen nicht nur in der Tätigkeit der Telomerase. Vielmehr erwiesen sich 21 andere Gene als bedeutsam, die an der DNA-Reparatur und an der Wartung der Telomere beteiligt sind. Diese Gene sind spezifisch bei Myotis myotis aktiv. Die Aktivität zweier Gene, ATM und SETX, ist besonders wichtig. Offenbar spielen also neben Telomerschutzprogrammen auch besondere Vorgänge in der DNA-Reparatur bei der erheblichen Verzögerung der Alterung eine Rolle.
Weshalb gerade das Große Mausohr der normalen Alterung entkommt, ist nicht bekannt. Jedenfalls zeigt dieses Beispiel, dass es bezüglich des Alterns sehr weit reichende Ausreißer geben kann. Vor diesem Hintergrund scheint auch ein sehr viel größeres individuelles Alter von Menschen nicht im Widerspruch zu biologischen Sachverhalten zu stehen.
[Fooley NM, Hughes GM et al. (2018) Growing old, yet staying young: The role of telomeres in bats’ exceptional longevity. Sci. Adv. 2018;4: eaao0926]
|
|
In Nordamerika von Alaska bis Kalifornien beheimatete Stahlkopfforellen (Oncorhynchus mykiss, Abb. 1) leben normalerweise sowohl in Süßwasser als auch zeitweise in Salzwasser und können ihre Osmoregulation entsprechend umschalten. Eine Population hat sich binnen 100 Jahren auf das Leben ausschließlich im Süßwasser spezialisiert. Die damit einhergehenden Änderungen des Erbguts erfolgten in kurzer Zeit, was nur auf der Basis einer ursprünglich großen Variationsbreite möglich war.
Die Fische schlüpfen in Süßwasser, verbleiben 1-2 Jahre dort, wandern dann über Flüsse in den pazifischen Ozean und verbringen dort 1-3 Jahre, um zur Laichzeit ins Süßwasser zurückzukehren.
 |
| Abb. 1: Stahlkopfforelle, auch als Regenbogenforelle bezeichnet. (National Digital Library of the United States Fish and Wildlife Service, gemeinfrei) |
|
Die Erfordernisse bezüglich der Osmoregulation (Gleichgewicht zwischen Salzen und Ionen innerhalb der Zellmembranen) sind im Süß- und Salzwasser gegensätzlich. Im Süßwasser müssen die Fische in der Lage sein, aktiv Ionen aus ihrer Umwelt aufzunehmen, um so den Salzverlust durch passive Diffusion auszugleichen, während im Salzwasser Ionen abgegeben werden müssen, um die Salzaufnahme durch ihren Körper auszugleichen. Fische, die in beiden Medien leben, benötigen also beide Mechanismen zur Aufrechterhaltung der erforderlichen Salzkonzentration in den Zellen und können zwischen ihnen umschalten. Darüber hinaus unterscheidet sich der Lebensraum im Süßwasser von der Salzwasserumgebung auch in der Wassertemperatur, in der Art der Strömung und in der Zusammensetzung der sonstigen Fauna.
Vor 120 Jahren wurden einige Stahlkopfforellen im Lake Michigan ausgesetzt. Nach anfänglichen Schwierigkeiten konnten die Fische eine stabile Population aufbauen. Für diese Fische war der große See gleichsam ein Ozean-Ersatz; sie wanderten nicht ins Meer, sondern verblieben dauerhaft im Süßwasser. Nun haben Forscher das Genom (komplettes Erbgut) von 264 Exemplaren der Stahlkopfforelle genetisch untersucht und eine Reihe von Unterschieden zwischen den Süßwasserformen und den in den Pazifik wandernden Formen entdeckt (Willoughby et al. 2018). Dabei zeigte sich, dass zwei veränderte Chromosomenregionen in einer direkten Verbindung mit der Osmoregulation stehen: Die Carboanhydrase (katalysiert die Hydratisierung von Kohlenstoffdioxid zu Kohlensäure und umgekehrt) und ein Transportprotein (SLC 26) sind in einer Weise verändert, dass die Ionenaufnahme aus dem Wasser erleichtert ist. Die oben erwähnte Fähigkeit, die Osmoregulation bei Süß- und Salzwasser umschalten zu können, ist energetisch kostspielig, sodass der Verlust dieser Fähigkeit bei dauerhafter Existenz im Süßwasser zudem von Vorteil ist (Willoughby et al. 2018, 8).
Die dritte veränderte Genregion betrifft das Enzym Ceramid-Kinase, das im Zusammenhang mit dem Stoffwechsel bei der Wundheilung der Tiere steht. Die im Süßwasser lebenden Tiere sind vermehrt Verletzungen durch Neunaugen ausgesetzt und Wunden heilen im Süßwasser schlechter; durch die Veränderung wird vermutlich der Heilungsprozess verbessert (Willoughby et al. 2018, 8).
Wie konnten sich die Gene aber so schnell verändern? Die Forscher fanden heraus, dass die Veränderungen weder auf Hybridisierungen mit anderen Stahlkopfforellen-Populationen noch auf Mutationen zurückgeführt werden konnten. Vielmehr sprechen die Befunde dafür, dass die anfangs überlebenden Individuen bereits die passenden Ausprägungen der relevanten Gene besaßen und daher am ehesten überleben konnten. Die spezialisierten Forellen des Lake Michigan „leben“ also von einer ursprünglichen genetischen Vielseitigkeit ihrer Vorläufer.
Bemerkenswert ist eine weitere Beobachtung: Die Wissenschaftler stellten fest, dass die genetische Vielfalt der Stahlkopfforellen im neuen Lebensraum in allen 29 Chromosomen deutlich niedriger liegt als bei den Populationen, die in den angestammten Gewässern leben, was auf einen Gründer-Effekt zurückgeführt wird. Spezialisierung ist wie in vielen anderen Fällen mit genetischer Verarmung verbunden.
Diskussion. Das Beispiel der Osmoregulation bei den Stahlkopfforellen reiht sich in eine lange Liste von Fällen ein, in denen von einer ursprünglich genetisch vielseitigen (polyvalenten) Ausgangssituation ausgegangen werden muss – soweit diese rekonstruiert werden kann – und in denen eine schnelle Veränderung beobachtet wurde (im Verlauf von Jahren bis Jahrzehnten). Die beobachteten Veränderungen und Spezialisierungen gehen mit Einschränkung der genetischen Vielfalt einher. Die ursprüngliche Vielseitigkeit kann enorm sein; immerhin ist es keine Kleinigkeit, sowohl im Süßwasser als auch im Salzwasser leben zu können.
Auf der Basis dieser Befunde kann man spekulieren, dass nach einer ursprünglich großen Toleranz bezüglich des Salzgehalts des Wassers bei heute weniger toleranten Arten sozusagen eine eingefrorene Spezialisierung erfolgt ist(verbunden mit genetischer Verarmung). Eine große Toleranzbreite könnte auch bei den umwälzenden Prozessen während der Sintflut eine wichtige Voraussetzung für das Überleben gewesen sein.
[Willoughby JR, Harder AM, Tennessen JA, Scribner KT & Christie MR (2018) Rapid genetic adaptation to a novel environment despite a genome-wide reduction in genetic diversity. Mol. Ecol., doi:10.1111/mec.14726]
|
|
Rote Blutzellen (Erythrozyten) sind bekanntlich verantwortlich für den Transport von Sauerstoff im Blut und für den Rücktransport von Kohlendioxid zu den Lungen. Diese Funktion basiert hauptsächlich auf der chemischen Struktur des roten Blutfarbstoffes Hämoglobin und den besonderen Eigenschaften ihres Zellskeletts. Die Abgabe von Sauerstoff beziehungsweise die Aufnahme von Kohlendioxid erfolgt in Kapillaren der Lungen bzw. des Zielgewebes; die Kapillaren sind die kleinsten Blutgefäße des Organismus. Sie sind etwa 0,5 mm lang und haben einen Durchmesser von 5 bis 10 µm. Der maximale Durchmesser der Erythrozyten beträgt ca. 7,5 µm und an ihren Rändern sind sie ca. 2 µm dick (Abb. 1). Diese frisbeeähnliche Form garantiert eine große Kontaktfläche für den Gasaustausch. Würde man die Oberflächen aller Erythrozyten nebeneinanderlegen, würde man bei einem gesunden Erwachsenen auf eine Fläche von ca. 4000-4500 m2 kommen, das entspricht etwa 75 % eines Fußballfeldes. Wie gelangen aber die 7,5 µm „dicken“ Blutzellen durch die teilweise nur 5 µm großen Kapillaren, ohne zu zerplatzen bzw. die Kapillaren zu verstopfen?
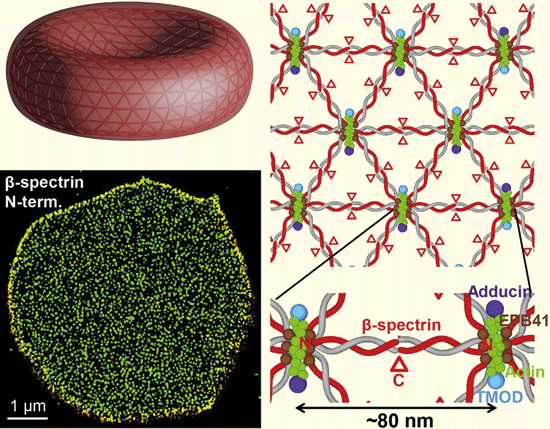 |
| Abb. 1: Oben links: Erythrozyt (rotes Blutkörperchen) mit seiner typischen frisbeeähnlichen Form, Durchmesser 7,5 µm, Dicke am Rand 2 µm. Unten links: Darstellung der für die Elastizität verantwortlichen Proteine (Spektrin) in der Membran von Erythrozyten (grüne Punkte), einzelne schwarze Lücken in dem Proteinteppich sind keine Defekte, sondern dienen einer schnellen Formänderung. Rechts: Schema der Anordnung der Proteinketten des Spektrins als Netzwerk, entspannt haben die einzelnen Knotenpunkte einen Abstand von 80 nm, unter Spannung kann der Abstand der Knotenpunkte bis auf 180-190 nm vergrößert werden. (Aus Pan et al. 2018, Abdruck mit freundlicher Genehmigung) |
|
Elektronenmikroskopisch konnte bislang an toten Erythrozyten gezeigt werden, dass ihre Membran (äußere Hülle) und ihr Zellskelett hochflexibel und sehr robust sind. Diese Untersuchungen beantworteten aber nicht alle Fragen. Eine neue innovative Mikroskopiertechnik mit einer örtlichen Auflösung von 25 nm (nm = Millionstel Millimeter) (STORM – Stochastic Optical Reconstruktion Microscopy) erlaubte es dem Team um L. Pan von der University of California in Berkeley, die Ultrastruktur des Zellskeletts lebender Blutzellen zu entschlüsseln. Dabei wiesen sie in den Membranen der Erythrozyten ein netzförmig aufgebautes Protein nach, das Spektrin, welches aus zu Dreiecken verknüpften, elastischen Molekülketten mit einer Kantenlänge von 80 nm besteht (Abb. 1b und c).
Vergleichbare netzförmige Proteinverbände wurden bereits in den Zellwänden von neuronalen Zellen (Zellen, die zum Nervensystem gehören) gefunden; dort zeigen die einzelnen Molekülketten eine Kantenlänge von 180-190 nm. Dieser Unterschied erklärt sich daraus, dass die Molekülketten bei Letzteren durch andere Zellbestandteile (Mikrotubuli, Neurofilamente) unter ständiger Spannung gehalten werden, die in den Erythrozyten fehlen, weshalb dort die Molekülketten entspannt bzw. relaxiert vorliegen. Das ist funktionell höchst sinnvoll.Die in den Erythrozyten angeordneten entspannten Molekülketten lassen eine Dehnung bis auf 190 nm zu und bilden damit ein hochflexibles federndes Maschenwerk. Daraus erklärt sich einerseits die hohe Verformbarkeit und Elastizität und anderseits die Robustheit der Erythrozyten, wenn sie durch die Kapillaren „gequetscht“ werden. Überrascht wurde die Forschergruppe durch den Befund, dass innerhalb des hochelastischen Netzwerkes große Lücken klaffen. Diese stellen keine Defekte und keine Fehlkonstruktion dar, sondern erlauben den Erythrozyten eine sehr schnelle Formänderung – so die Interpretation der Forscher. Die Löcher wirken hier wie Beugepunkte und garantieren ein noch größeres Potenzial für eine kurzfristig erforderliche Verformbarkeit in den Kapillaren. Diese Erkenntnisse sind auch für die medizinische Erforschung der Malaria von größtem Interesse, da bekannt ist, dass die Malaria-Erreger mit dem Zellskelett der roten Blutkörperchen interagieren. Blutzellen, die ihre Flexibilität und damit die Funktionalität des oben beschriebenen elastischen Netzwerkes der Zellmembran aufgrund von Veränderungen der chemischen Struktur des Hämoglobins verloren haben (sogenannte Sichelzellen), sind bekanntlich resistent gegenüber den Malariaerregern.
Diskussion. Der von der Arbeitsgruppe um L. Pan beschriebene Merkmalskomplex ist ein herausforderndes Design-Merkmal. Das elastische maschenartige Netzwerk ist nicht genetisch codiert (nur dessen Eiweißbausteine), aber höchst präzise in die physiologischen Wechselwirkungen sowie anatomischen Gegebenheiten eingepasst, und das über mehrere hierarchische Stufen, von der molekularen Ebene im Nanometerbereich bis zur Organebene. Ein evolutionärer Mechanismus, der unabhängig von Veränderungen der genetischen Grundlagen kreativ agiert und, wie hier gezeigt, die beste stereochemische Konfiguration von Proteinen in Korrelation zu den lokal erforderlichen funktionellen Anforderungen ermöglicht, ist unbekannt. Rote Blutzellen sind Abkömmlinge von Stammzellen der Blutbildung, in denen ein riesiges Potential an Entwicklungsprogrammen angelegt ist (z. B. für die bekannte Vielzahl weißer Blutzellen oder für Thrombozyten). Diese Entwicklungsprogramme werden unter dem Einfluss hochkomplexer biochemischer Steuermechanismen aktiviert oder unterdrückt. Die evolutionäre Ableitung roter Blutkörperchen und des speziell hier beschriebenen elastischen Netzwerkes innerhalb der Zellmembranen ist ohne koordinierte und korrelierende Veränderungen des ursprünglichen Blutbildungssystems undenkbar und deshalb auf der Basis bisher diskutierter Evolutionsmechanismen nicht einmal in Ansätzen zu verstehen.
[Pan L, Yan R, Li W & Ku K (2018) Super-Resolution Microscopy Reveals the Native Ultrastructure of the Erythrocyte Cytoskeleton. Cell Reports 22, 1151-1158.]
|
|
|
|
