 |
 Studium Integrale Journal
Studium Integrale Journal
23. Jahrgang / Heft 1 - Mai 2016
Seite 55 - 65
|
| |
|
Vor knapp 60 Jahren schlugen die beiden bekannten Biologen W. L. Brown und E. O. Wilson die evolutionsbiologische Hypothese des „character displacement“ vor. Diese sagt voraus, dass zwei eng verwandte Spezies, die beide in einem bestimmten Gebiet leben, zueinander in Konkurrenz treten werden. Um diese Konkurrenz zu vermeiden, würden sie sich auseinanderentwickeln – eine Ursache von evolutionärer Divergenz (Brown & Wilson 1956).
Nun liegt eine Arbeit einer Gruppe von Wissenschaftlern vor, die diese Hypothese in einem Freilandexperiment mit Anolis-Eidechsen testeten (Stuart et al. 2014). Anolis ist eine Gattung von Eidechsen, die im karibischen Raum häufig vorkommt und ziemlich gut erforscht ist. Da sie eine relativ kurze Generationszeit hat (unter einem Jahr) und ihr Verbreitungsgebiet viele kleine Inseln umfasst, eignet sie sich sehr gut für solche Experimente (vgl. Vedder 2012).
In diesem Fall war das Hauptuntersuchungsobjekt A. carolinensis, eine Art, die in den südlichen USA beheimatet ist (Abb. 1). Wie viele ihrer Verwandten leben Eidechsen dieser Art auf Sträuchern und Bäumen. Gibt es keine anderen Anolis-Arten in der Umgebung, so verbreiten sie sich über die gesamte Höhe des Baumes. Es wurde jedoch beobachtet, dass in Gegenden, wo A. carolinensis gemeinsam mit A. sagrei (Abb. 2) vorkommt, A. carolinensis meistens höher oben im Geäst sitzt als A. sagrei. Die Forscher vermuten, dass dieses Verhalten ausgelöst wird, um die interspezifische Konkurrenz zwischen A. carolinensis und A. sagrei zu vermindern. Wenn dem so wäre, würde das die „character displacement“-Hypothese stützen.
 |
| Abb. 1: Rotkehlanolis (Anolis carolinensis). Männchen mit aufgestelltem Kehllappen. (CC BY-SA 3.0) |
|
1995 begannen die Forscher mit einem Experiment, in dem sie sechs kleine Inseln vor der Küste Floridas untersuchten. Die Inseln beherbergten zu der Zeit nur A. carolinensis. Die Wissenschaftler ermittelten ihre durchschnittliche Sitzhöhe. Daraufhin setzten sie auf drei der Inseln A. sagrei-Eidechsen aus. In den nächsten drei Jahren beobachteten sie, dass sich wie vorhergesagt die durchschnittliche Sitzhöhe von A. carolinensis auf den Inseln mit zwei Spezies etwa verdoppelte, während sie auf den Inseln ohne A. sagrei konstant blieb.
 |
| Abb. 2: Bahama-Anolis (Anolis sagrei) in Florida. (Foto: Ianaré Sévi, gemeinfrei) |
|
15 Jahre später kehrten sie zu der Inselgruppe zurück, um weitere Untersuchungen vorzunehmen. Diesmal vermaßen sie die Zehen der Eidechsen. Ähnlich wie Geckos weist auch A. carolinensis Zehenpolster mit Lamellen (kleine Hautfalten, die die Oberfläche und Bodenhaftung vergrößern) auf. Es ist bekannt, das größere Zehenpolster mit mehr Lamellen beim Klettern auf dünnen Ästen einen Vorteil bieten. Folglich erwarteten die Autoren, dass die höher lebenden A. carolinensis-Eidechsen auf den Inseln mit A. sagrei diese Anpassung aufweisen sollten, eine Erwartung, die sie wiederum bestätigen konnten. Auf den Inseln ohne A. sagrei blieb auch in dieser Hinsicht alles beim Alten. Außerdem konnten die Wissenschaftler bestätigen, dass es sich bei diesem Phänomen tatsächlich um parallele genetische Adaption handelt und es nicht etwa durch Plastizität (umweltbedingte Anpassung) oder Migration (Einwanderung aus anderen Gebieten) hervorgerufen wurde.
Die Bedeutung dieser Studie liegt zum einen in der Geschwindigkeit, mit der A. carolinensis sich an die neuen Gegebenheiten anpasste. Innerhalb von zwei Monaten nach der Invasion von A. sagrei ließ sich bereits eine verhaltensbiologische Adaption beobachten (Verschiebung der Sitzhöhe) und nach weniger als 20 Generationen gab es genetisch bedingte morphologische Änderungen (die Vergrößerung der Zehenpolster). Daher zeigt die Studie auch, dass sich manche evolutionsbiologische Fragestellungen in verhältnismäßig kurzer Zeit untersuchen lassen. Es braucht keine Jahrmillionen, um Evolution in Form von genetischer Anpassung in der Natur beobachten zu können. Zum anderen stützen die Ergebnisse die „character displacement“-Hypothese sehr gut. Browns und Wilsons Hypothese kann also als ein bestätigter Mechanismus der (Mikro-)Evolution angesehen werden.
[Brown WL & Wilson EO (1956) Character Displacement. Syst. Zool. 5, 49-64 • Stuart YE, Campbell TS et al. (2014) Rapid evolution of a native species following invasion by a congener. Science 346, 463-466 • Vedder D (2012) Gründereffekt bei Eidechsen: ein Freilandexperiment auf Inseln. Stud. Integr. J. 19, 107-109]
|

|
Die fossile Überlieferung der Vögel zeigt zwei Phasen eines plötzlichen Erscheinens vielfältiger Formen. Die eine liegt am Beginn des Tertiärs (wo die auch heute vorkommenden Vogelordnungen explosiv im Fossilbericht auftauchen), die andere in der Unterkreide, in deren geologischen Schichten ebenfalls ziemlich abrupt eine große Vielfalt von Vögeln mit anderer Anatomie als der heutigen überliefert ist. Der berühmte „Urvogel“ Archaeopteryx aus dem Oberjura datiert noch ca. 20 Millionen radiometrische Jahre älter; es ist aber nach wie vor umstritten, ob er aktiv fliegen konnte oder eher ein Gleiter war. Andere eindeutig befiederte Formen aus dem Oberjura (z. B. Anchiornis) gelten als flugunfähig.
Die Kreidevögel unterscheiden sich anatomisch mehr oder weniger deutlich von den tertiären Vögeln und zeigen manche Gemeinsamkeiten mit zeitgleich lebenden Theropoden-Dinosauriern (zweibeinige Raubdinosaurier), die als ihre stammesgeschichtlichen Vorfahren gelten. Sie werden in zwei Gruppen unterteilt, die Enantiornithes („Gegenvögel“; sind am Ende der Kreide ausgestorben) und die Ornithurae („Vogel-Schwänze“), zu denen u. a. auch die heutigen Vögel gehören.(Es gibt in der Fachliteratur keine einheitliche Begründung für die Bezeichnung „Gegenvögel“. Es wird auf zwei anatomische Besonderheiten hingewiesen, die die Enantiornithes von anderen Vögeln unterscheiden: zum einen die Art der Verwachsung einiger Fußknochen, zum anderen der Bau des Gelenks zwischen Schulterblatt und Rabenbein. Bei den Gegenvögeln sind Gelenkkopf und Gelenkpfanne im Vergleich zu allen anderen Vögeln sozusagen vertauscht. Es ist schwer denkbar, wie die eine anatomische Ausprägung in die andere evolutiv überführt werden könnte. Für die Ornithurae gibt es keine eingebürgerte deutsche Bezeichnung; der Name bezieht sich auf den kurzen knöchernen Schwanz, wie er auch für die heutigen Vögel typisch ist.)
Eine Forschergruppe hat nun herausgefunden, dass die Enantiornithes trotz deutlich verschiedener Anatomie im Vergleich zu heutigen Vögeln überraschenderweise vergleichbare Fähigkeiten zu einem versierten Flug besaßen. Nachgewiesen wurde dies anhand eines sehr gut erhaltenen vorderen Teils einer Vorderextremität aus einem Kalkstein von Las Hoyas, Spanien (datiert auf 125 Millionen radiometrische Jahre), die aufgrund ihres Baus von einem enantiornithinen Vogel stammt (Navalón et al. 2015; Abb. 1). Erhalten sind auch Hand- und Armschwingen. Das Besondere an dem Fund: Es sind Details von Muskeln, Bändern und Follikeln (in der Haut befindliche Basis der Federn) aus den Bereichen zwischen den Flügelteilen und Fingern (Propatagium, Postpatagium und Patagium der Alula) in Form von Abdrücken erhalten (Die Alula = Daumenfittich ist wichtig z. B. für Manöver beim Landen). Der Fund aus Las Hoyas lässt auch Verbindungen von Bindegewebsstrukturen mit den Ansätzen der Flugfedern erkennen; das gesamte komplizierte Netzwerk weist große Ähnlichkeiten mit den anatomischen Verhältnissen heutiger Vögel auf. Ein solch komplexes System aus Sehnen, Bändern, Bindegewebe und Muskeln ermöglicht eine ausgefeilte Manövrierbarkeit während des Fluges.
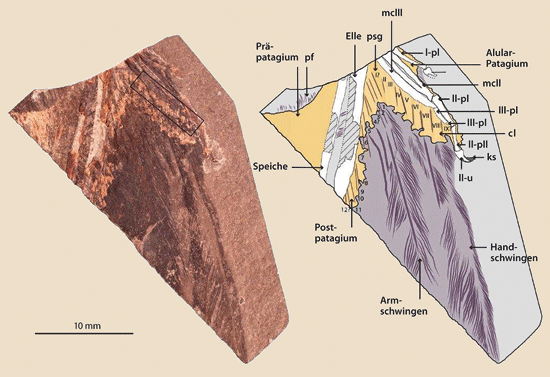 |
| Abb. 1: Fossil und intepretierende Zeichnung des Enantiornithinen MCCMLH31444 aus Las Hoyas, Spanien. Der umrahmte Teil umfasst die Region des Übergangs zwischen Knochen und Weichgewebe. Abkürzungen: I, II, III bezieht sich auf Finger I, II und III, cl calamus; ks keratinisierte Hülle, mc Metacarpale, p Fingerknochen, pf Daunenfedern, u Klaue. (Aus Navalón, 2015, verändert) |
|
Aus ihren Beobachtungen schließen die Autoren, dass die „Gegenvögel“ der Kreidezeit ebenso gut wie viele moderne Vögel „über den Köpfen der Dinosaurier“ fliegen konnten, wie der Leiter der Forschergruppe, L. M. Chiappe, es formuliert (http://www.sciencenutshell.com/the-bird-who-flew-above-dinosaurs-heads/). „The anatomical match between the muscle network preserved in the fossil and those that characterize the wings of living birds strongly indicates that some of the earliest birds were capable of aerodynamic prowess like many present-day birds“ (Chiappe unter http://www.eurekalert.org/pub_releases/2015-10/nhmo-taf100615.php). Bislang war man davon ausgegangen, dass die Gegenvögel der Unterkreide eher schlechte Flieger waren; das kann in dieser Pauschalität mit diesem Fund als widerlegt gelten. Die Gegenvögel starten gleichsam als gut ausgebildete Flieger; eine Evolution der Flugfähigkeit ist innerhalb dieser Gruppe somit nicht dokumentiert. Das Beispiel zeigt auch, dass es zu Fehlschlüssen kommen kann, wenn man die Funktionalität nur anhand von Knochenüberresten erschließen möchte (bzw. kann).
Navalón et al. (2015) stellen allgemein über die Gegenvögel am Schluss ihres Artikels fest, dass obwohl die Enantiornithinen eine Anzahl von primitiven Skelettelementen aufwiesen, bereits die frühesten unter ihnen (Protopteryx fengningensis) Vorderextremitäten mit modernen Proportionen besaßen, außerdem ein gekieltes Brustbein und einen „fortschrittlichen“ Schultergürtel mit einem Kanal für die Passage der Flugmuskeln (triosseal canal), was alles auf die Fähigkeit zu aktivem Flug und Flügelschlag ähnlich dem heutiger Vögel hinweise. Auch Ähnlichkeiten der Körperbedeckung mit heutigen Vögeln mit identischer Federanordnung würden in diese Richtung weisen.
Es sei noch angemerkt, dass auch unter den Ornithurae eine der ältesten Formen als guter Flieger angesehen wird (Wang et al. 2015).
[Navalón G, Marugán-Lobón J, Chiappe LM, Sanz JL & Buscalioni A (2015) Soft-tissue and dermal arrangement in the wing of an Early Cretaceous bird: Implications for the evolution of avian flight. Sci. Rep. 5:14864 • Wang M et al. (2015) The oldest record of ornithuromorpha from the early cretaceous of China. Nat. Comm. 6:6987, doi: 10.1038/ncomms7987]
|
|
|
Seit der Erstveröffentlichung von Fundmaterial von Homo habilis im Jahre 1964 gibt es eine kontroverse Diskussion um die Gültigkeit dieses Taxons: War habilis wirklich ein Mensch oder gehört das Fundmaterial eher zu den großaffenähnlichen Australopithecinen?
Zachwieja et al. (2016) haben einen wichtigen Beitrag zu dieser Kontroverse geleistet, indem sie das Verhältnis der Knochenstärke des Oberarmes (Humerus) zum Oberschenkel (Femur) der fossilen Homininen (= Menschen und hypothetische evolutionäre Vorläufer des Menschen) KNM-WT 15000 (Homo erectus), AL 288-1 (Australopithecus afarensis) und OH 62 (Homo habilis; Abb. 1) im Vergleich mit heute lebenden Menschen und nichtmenschlichen Primaten bestimmten. Dieses Verhältnis – der humerofemorale Index – spiegelt die Belastung der Extremitäten bei der Fortbewegung wider.
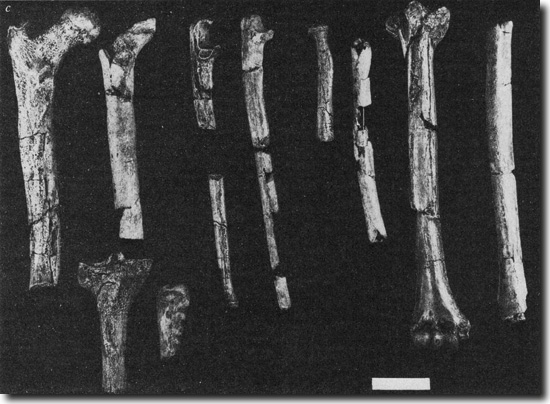 |
| Abb. 1: Langknochen von OH 62 (Homo hablilis) im Vergleich mit entsprechenden Elementen von AL 288-1 (Australopithecus afarensis); die Knochen von OH 62 sind jeweils rechts. Balken: 4 cm. (Aus Johanson et al. 1987) |
|
Die Untersuchung ergab einen humerofemoralen Index für KNM-WT 15000 im Bereich des modernen Menschen. Dagegen liegen AL 288-1 und OH 62 im Variationsbereich des Schimpansen. Nach diesem Ergebnis kletterten Homo habilis und Australopithecus afarensis – im Gegensatz zu Homo erectus – gewohnheitsmäßig in Bäumen. Zachwieja et al. (2016) bestätigen mit diesem Resultat das Ergebnis einer ähnlichen Untersuchung von Ruff aus dem Jahre 2009.
Mit der heute in der Paläanthropologie leider häufigen Zuordnung des habilis-Materials zum Menschen umfasst die Gattung Homo einerseits Arten wie erectus mit einem modern-menschlichen Extremitätenbelastungsmuster und andererseits habilis mit einer schimpansenähnlichen Belastung wie die Australopithecinen. Dagegen werden beispielsweise Gorilla und Schimpanse, deren Fortbewegung zwar nicht identisch (Schmitt et al. 2016), aber doch relativ ähnlich ist, da beide einen Knöchelgang praktizieren, in unterschiedliche Gattungen gestellt. Aufgrund der Australopithecus-ähnlichen Fortbewegung und vieler weiterer nichtmenschlicher Merkmale ist die Zuordnung von habilis zum Formenkreis der großaffenähnlichen Australopithecinen viel begründeter als eine Zuordnung zum Menschen.
[Johanson DC, Masao FT et al. (1987) New partial skeleton of Homo habilis from Olduvai Gorge, Tanzania. Nature 327, 205-209 • Ruff C (2009) Relative limb strength and locomotion in Homo habilis. Amer. J. Phys. Anthropol. 138, 90-100 • Schmitt D, Zeininger A, Hamrick E, Snyder ML, Kivell TL & Wunderlich RE (2016) Gorilla limb kinematics and hominoid locomotor diversity: Implications for hominin locomotor evolution. Amer. J. Phys. Anthropol. 159, S62, 282 • Zachwieja AJ, Demes B, Jungers WL, Carlson KJ, Grine FE, Pearson OM, Shackelford LL & Polk JD (2016) Ratios of humeral to femoral mid-shaft cortical area reflect differences in locomotor behavior in primates, including fossil hominins. Amer. J. Phys. Anthropol. 159, S62, 342-343.]
|
|
In der Paläanthropologie ist es üblich, bei fossilen Menschen und deren angenommenen Vorläufern (Hominine) Unterschiede in der absoluten Größe des Gehirns und der relativen Größe von Gehirnarealen im Rahmen eines makroevolutionären Prozesses funktionell zu deuten. Das heißt: Unterschiede werden als Indizien für unterschiedliche evolutionäre Entwicklungsstadien interpretiert. Eine jüngere Untersuchung verdeutlicht jedoch die Schwierigkeit dieses Unterfangens.
Albessard et al. (2016) haben die Gehirne von Menschen aus dem Jungpaläolithikum von Cro-Magnon und Abri Pataud (Frankreich), dem Epipaläolithikum von Afalou (Algerien) und Taforalt (Marokko) und aus der heutigen Zeit analysiert. Im Evolutionsmodell sind diese Menschen phylogenetisch gleichwertig.
Die Untersuchung ergab, dass sich das menschliche Gehirn in den vergangenen 20.000 radiometrischen Jahren verkleinerte und verkürzte. Außerdem kam es zu Änderungen der relativen Größe von Gehirnarealen: Die Stirn- und Hinterhauptlappen wurden kleiner, während sich die Scheitellappen und das Kleinhirn relativ vergrößerten.
Diese hohe Plastizität des menschlichen Gehirns macht es nach Albessard et al. (2016) problematisch, die Größe und die Form fossiler Gehirne funktionell zu interpretieren.
[Albessard L, Durrleman S, Balzeau A & Grimaud-Hervé D (2016) The human brain evolving: a diachronic study of endocranial variation. Amer. J. Phys. Anthropol. 159, S62, 76-77.]
|
|
Es ist der Normalfall, dass die Fossilüberlieferung diskontinuierlich ist. In einem evolutiven Szenario wechseln demnach Phasen von weitgehendem Stillstand mit dem relativ plötzlichen Erscheinen zahlreicher neuer Formengruppen ab. An der bereits von Darwin beschriebenen Situation hat sich im Großen und Ganzen bis heute nichts geändert, auch wenn zahlreiche Mosaikformen gefunden werden, die z. T. als Übergangsformen zwischen verschiedenen Großgruppen von Lebewesen interpretierbar sind. Eine auffällige Diskontinuität zeigt sich auch bei einer der berühmtesten Fossilgruppen, den Dinosauriern. Vor einigen Jahren stellten Martinez et al. (2011) fest, dass die fossile Überlieferung der drei Hauptgruppen der Dinosaurier etwa gleichzeitig in der Obertrias beginnt (unter den Saurischiern [Echsenbeckensaurier] sind das die Theropoden und Sauropoden, außerdem sind auch die Ornithischier [Vogelbeckensaurier] nachgewiesen). Die frühen Dinosaurier seien sowohl verbreiteter als auch vielfältiger gewesen als zuvor angenommen. Seltsamerweise habe es dann aber noch 30 Millionen Jahre gedauert, bis sie die Fauna dominierten.
Die Dinosaurier werden evolutionär von einer Gruppe von Echsen abgeleitet, die als Dinosauromorpha bezeichnet werden; dieser Name soll ausdrücken, dass diese Formen den Dinosauriern im Körperbau relativ nahe stehen, aber keine Dinosaurier sind. Die Fundschichten der Dinosauromorpha wurden bislang der Mittleren Trias zugeordnet. Grundlage für diese Zuordnung waren Faunenvergesellschaftungen, die für diese Schichten als typisch gelten. Radiometrische Datierungen lagen dagegen bis vor kurzem noch nicht vor. Dinosaurier-Gruppen wurden bisher in Schichten fossil gefunden, die nur etwa 10 Millionen radiometrische Jahre jünger datiert wurden. Diese (im Langzeitrahmen) kurze Zeitspanne galt schon bisher in evolutionstheoretischer Sicht als Beleg für eine überraschend schnelle Entstehung der Dinosaurier.
Nun konnte eine Forschergruppe (Marsicano et al. 2016) mit Hilfe der Uran-Blei-Methode Zirkone datieren, die in der triassischen Chañares-Formation (Argentinien) eingelagert sind, welche Dinosauromorpha-Fossilien enthalten, und erlebte eine Überraschung: Das radiometrische Alter erwies sich als etwa 5-10 Millionen Jahre geringer als bisher angenommen, so dass die Chañares-Formation nun in die Obertrias gestellt werden muss (bisher Mitteltrias, s. o.; auf 236-234 Millionen radiometrische Jahre datiert). Auch die ältesten dinosaurier-führenden Schichten konnten mit dieser Methode datiert werden und erhielten ebenfalls ein jüngeres Alter (231 Millionen radiometrische Jahre). Folge: Der ohnehin geringe zeitliche Abstand zwischen den Dinosauromorpha und den ältesten fossil nachweisbaren Dinosauriergruppen ist noch kleiner und beträgt weniger als 5 Millionen radiometrische Jahre – sehr wenig Zeit (zu wenig?) für eine evolutionäre Entstehung. Die Ergebnisse der Datierungen können sehr wahrscheinlich auch auf die anderen Fundorte mit Dinosauromorpha übertragen werden, die bislang noch nicht radiometrisch datiert werden konnten – so die Autoren.
 |
| Abb. 1: Skelett eines Lagosuchus talampayensis; einer früh fossil überlieferten Dinosaurier-Gattung aus der Mitteltrias Argentiniens. (CC BY-SA 3.0) |
|
Eine weitere Folge der Neudatierung ist, dass die bisherige Erklärung für das Aufkommen der Dinosaurier fallengelassen werden muss, wonach die Dinosaurier nach einem großen Aussterbeereignis an der Perm-Trias-Grenze die frei gewordenen Lebensräume füllen konnten. Dafür aber treten die Dinosaurier – gemäß der fossilen Überlieferung – nämlich viel zu spät auf. Davon abgesehen machen freie ökologische Räume ohnehin noch keine neuen Baupläne. Wahrscheinlicher ist bei einem relativ plötzlichen Erscheinen neuer Formen das Heraustreten aus bisher geologisch nicht überlieferten Lebensräumen, also die zahlenmäßig deutliche Zunahme von zuvor bereits existenten, aber fossil nicht überlieferten Formen (vgl. Stephan 2015, 129f., 157).
[Marsicano CA, Irmis RB, Mancuso AC, Mundile R & Chemale F (2016) The precise temporal calibration of dinosaur origins. Proc. Natl. Acad. Sci. 113, 509-513 • Martinez RN, Sereno P, Alcober OA, Colombi CE, Renne PR, Montanez IP & Currie BS (2011) A basal dinosaur from the dawn of the dinosaur era in southwestern Pangaea. Science 331, 206-210 • Stephan M (2015) Sintflut und Geologie. Holzgerlingen, 4. Aufl.]
|
|
Im Jahr 2013 veröffentlichten zwei ausgewiesene Kenner, Douglas Erwin und James Valentine, das Buch „The Cambrian Explosion“. Der Titel bezieht sich auf die Art und Weise, wie sich die Fossilüberlieferung im unteren Teil des Kambriums darstellt: Zahlreiche grundverschiedene Tierbaupläne tauchen erstmals auf, und zwar in großer Fülle und Verschiedenartigkeit und zudem in weiter geographischer Verbreitung. Erwin & Valentine (2013, v) schreiben: „… während einer relativ kurzen Zeitspanne im frühen Kambrium erfolgt ein explosives Erscheinen vieler verschiedener, gestaltlich abgegrenzter Fossilien, einschließlich Vertreter der meisten größeren Tiergruppen, die auch heute leben“ (vgl. Abb. 2).
 |
| Abb. 1: Rekonstruierte Szenerie des kambrischen Meeresbodens mit dem räuberisch lebenden Anomalocaris. (© John Sibbick/Natural History Museum) |
|
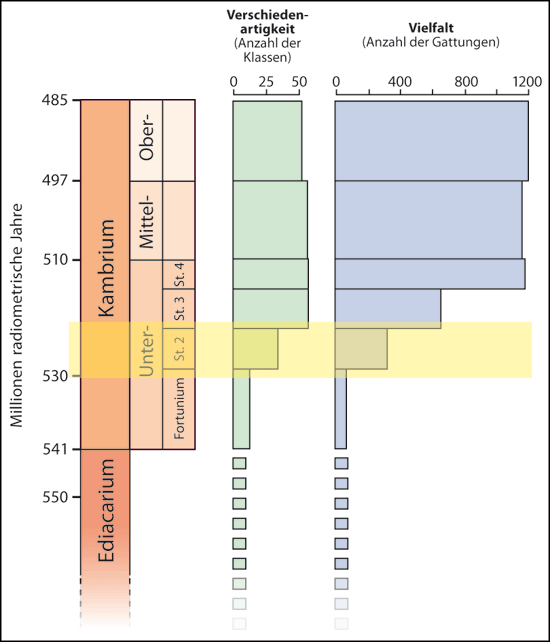 |
| Abb. 2: Kambrische Explosion, dargestellt anhand der Anzahl der Klassen (Verschiedenartigkeit; Mitte) und der Anzahl der Gattungen (Vielfalt; rechts). Im oberen Teil des Unterkambriums (farblich unterlegt) erfolgt eine rasante Zunahme. St.=Stufe. (Nach Marshall 2006, vereinfacht) |
|
So klar der fossile Befund ist, so unklar ist, wie er im evolutionstheoretischen Rahmen erklärt werden könnte. Was war der Auslöser für diesen „evolutionären Durchbruch“ (Fox 2016)? In neuen Untersuchungen wird der schon älteren Idee nachgegangen, dass ein Anstieg des Sauerstoffgehaltes ausschlaggebend gewesen sein könnte. Schließlich sind Innovationen wie Muskeln, Nervensystem, Einrichtungen zum Beuteerwerb und zur Verteidigung, mineralisierte Schalen und Skelettelemente sehr energiehungrig und damit auch sauerstoffbedürftig. Die gegenüber den Formen des nächst älteren Ediacariums neu vorkommende räuberische Lebensweise erforderte größere Mengen an Sauerstoff, sie habe zudem zu einem Wettrüsten zwischen Räubern und Beute geführt (Fox 2016, 269), was ebenfalls die Evolution von Neuheiten begünstigt habe.
Um die Idee mit dem Sauerstoffgehalt zu prüfen, wurden Gesteine, die ehemals am Meeresgrund entstanden sind und dem Kambrium oder älteren Sedimenten zugeordnet werden, auf das Vorkommen von Eisen, Molybdän und anderen Metallen untersucht (Sperling et al. 2015). Die Löslichkeit dieser Metalle hängt eng mit der Menge an Sauerstoff zusammen, so dass diese Metalle als Indikatoren dafür genutzt werden können, wie viel Sauerstoff im Wasser war, als die betreffenden Sedimente gebildet wurden.
Erik Sperling von der Stanford University in Kalifornien und sein Team sammelten Daten von 4.700 Eisenmessungen weltweit aus Proben aus dem Kambrium und dem Ediacarium – und fanden keinen besonders auffälligen Sauerstoff-Anstieg an der (künstlichen) Grenze zwischen den beiden geologischen Perioden. Der Sauerstoffgehalt heutiger Meere sei im Kambrium nicht erreicht worden. Andererseits weiß man von heutigen sauerstoffarmen Ökosystemen im Meer, dass auch bei einem sehr viel niedrigeren Sauerstoffgehalt als dem gegenwärtigen komplexes Leben möglich ist. Daher könnte – so Sperling – auch ein moderater Anstieg des Sauerstoffgehalts als Auslöser für eine „große Veränderung“ ausgereicht haben (Fox 2016, 269).
So umfangreich und detailliert die neuen Untersuchungen sind – welcher Sachverhalt wird damit eigentlich getestet? Offenkundig wird damit nicht eine Hypothese darüber geprüft, wie die Fülle der Tierbaupläne entstanden ist! Nachgewiesen wird bestenfalls ein Zusammenhang zwischen dem Sauerstoffgehalt in den Meeren und der Komplexität der in den betreffenden Sedimenten erhaltenen Lebewesen und Lebensgemeinschaften. Mit Evolution hat das nichts zu tun. Der festgestellte Zusammenhang ist ein ökologischer, kein evolutionärer. Wenn man dennoch einen evolutionären Kontext voraussetzt, könnte man auch sagen, dass auf eine mögliche Begleiterscheinung bzw. eine Koinzidenz oder eine notwendige Bedingung der kambrischen Explosion getestet wurde. Eine Koinzidenz ist aber keine Ursache. Schließlich ist der heutige Sauerstoffgehalt der Luft auch keine Ursache für die Entstehung eines Verbrennungsmotors, sondern nur eine notwendige Voraussetzung. Dieser selbstverständliche Sachverhalt wird immer wieder verschleiert, indem nicht klar zwischen Ursachen und notwendigen Voraussetzungen unterschieden wird. Kurz: Auslöser machen keine Baupläne.
Eine weitere aktuelle Arbeit zum Thema soll hier noch Erwähnung finden – eine Studie, von der die Neue Zürcher Zeitung (NZZ) behauptet, dass sie einen „ziemlich grossen Nagel in den Sarg der ‚kambrischen Explosion‘“ schlagen würde (www.nzz.ch/wissenschaft/biologie/jahrmillionen-mit-der-dna-zurueckgeblickt-1.18660890). Es geht in der betreffenden Arbeit um die Ermittlung der Divergenzzeiten der Tierstämme, die im Kambrium fossil plötzlich in Erscheinung treten. Diese (hypothetischen) Zeiten werden durch Anwendung der sogenannten Molekularen Uhr ermittelt. Diese wiederum beruht auf einem Vergleich von DNA-Sequenzen und einer Kalibrierung mit Hilfe geeigneter Fossilien. Unter evolutionstheoretischen Voraussetzungen – also nicht alleine anhand der Sequenzdaten und Fossilfunde – kann auf eine Ganggeschwindigkeit des evolutiven Wandels geschlossen werden, und damit wiederum können Aufspaltungszeitpunkte verschiedener Linien von Organismengruppen bestimmt werden. In der Vergangenheit hatten verschiedene Studien sehr unterschiedliche Zeiten z. B. für die Existenz des letzten gemeinsamen Vorfahren aller Bilaterier (Zweiseitentiere) ergeben, wobei sich ein Trend abzeichnete, dass dieser hypothetische gemeinsame Vorfahr mindestens 100 Millionen radiometrische Jahre vor der kambrischen Explosion existierte. Die neue Studie, auf die sich die NZZ bezieht, bringt diesbezüglich nichts grundsätzlich Neues; sie ist jedoch sehr viel detaillierter und differenzierter als bisherige Arbeiten (Reis et al. 2015). Ein Ergebnis dieser Arbeit ist, dass – Evolution vorausgesetzt – die kambrische Explosion ein Erhaltungsartefakt sein müsste – sprich: Die kambrischen Tierstämme sollten bereits vor 833-650 Millionen radiometrischen Jahren entstanden sein (eine genauere Bestimmung geben die Daten trotz aller Fülle nicht her); entsprechende Fossilien werden aber viel später gefunden. Folgt man dieser Deutung der molekularen Daten, ergibt sich das Problem, warum die Fossilisierung so stark verspätet und eben damit doch plötzlich eintritt. So oder so: Der Fossilbericht bezeugt ein abruptes Erscheinen vielgestaltiger Organismengruppen und voll funktionsfähiger Organismen (die „Explosion“); daran ändern die neuen Daten nichts. Erschwerend kommt hinzu, dass die neuen Daten kaum auf evolutionäre Aufzweigungsabfolgen schließen lassen, so dass die Autoren es für verfrüht ansehen, evolutionäre Abfolgen der kambrischen Tierstämme auf der Basis der molekularen Uhr zu rekonstruieren. Nebenbei: Wenn die kambrischen Stämme wirklich vor mindestens 650 Millionen radiometrischen Jahren entstanden wären, wären alle Überlegungen zu Sauerstoffgehalten im Ediacarium und Kambrium bedeutungslos.
[Erwin DH & Valentine JW (2013) The Cambrian explosion. Austin: Roberts & Company Publishers • Fox D (2016) What sparked the Cambrian explosion? Nature 530, 528-530 • Reis DM, Thawornwattana Y, Angelis K, Telford MJ, Donoghue PC & Yang Z (2015) Uncertainty in the timing of origin of animals and the limits of precision in molecular timescales. Curr Biol. 2015, doi: 10.1016/j.cub.2015.09.066 • Sperling EA, Wolock CJ et al. (2015) Statistical analysis of iron geochemical data suggests limited late Proterozoic oxygenation. Nature 523, 451-454.]
|
|
Die Venusfliegenfalle (Dionaea muscipula) ist eine der faszinierendsten Pflanzen – sie ist bekannt durch ihre ungewöhnliche karnivore (fleischfressende) Lebensweise. Da die Pflanze auf nährstoffarmen Böden wächst, ist sie auf eine zusätzliche Nährstoffquelle angewiesen. Ein wichtiger Mineralstoff für die Pflanze ist Natrium. Um an diesen Stoff zu gelangen, lockt sie mit zwei roten Blattspreiten, etwas Nektar und einem UV-Muster deshalb ihre Beute an. Lässt sich ein Insekt auf einer der Blattspreiten nieder, berührt es oft die sich darauf befindenden Fühlborsten. Durch jede Berührung mit einer Fühlborste wird ein Aktionspotential generiert. Falls auf das erste Aktionspotential innerhalb von 10-15 Sekunden ein zweites folgt, schnappen die beiden Blattspreiten in einem Bruchteil einer Sekunde zusammen. Die Falle „zählt“ also auf zwei.
 |
| Abb. 1: Venusfliegenfalle (Dionaea muscipula). (Joe MiGo, CC BY-SA 3.0) |
|
 |
| Abb. 2: Einzelnes Fangblatt mit den Fühlborsten. (CC BY-SA 2.5) |
|
Es wird vermutet, dass der grüne Jäger nicht bereits nach der ersten Berührung mit einer Fühlborste zuschnappt, da ein Fehlalarm vorliegen könnte. Dies würde für die Pflanze einen großen Energieverlust bedeuten, da die Falle vergebens zuschnappt und sich umsonst wieder öffnen muss. Die Falle hat also einen Sicherungsmechanismus.
Längliche Borsten am Rand beider Blattspreiten, welche in geschlossenem Zustand wie Finger von gefalteten Händen ineinandergreifen (Abb. 1), verhindern das Entkommen von lukrativer größerer Beute. Im dadurch geformten „grünen Magen“ wird das Tier mit Hilfe eines Enzymcocktails verdaut und die daraus gewonnenen Nährstoffe werden von der Pflanze aufgenommen.
Ein Forschungsteam der Universität in Würzburg (Böhm et al. 2016) konnte vor Kurzem zeigen, dass die Falle sogar weiter zählen kann. Wird eine Fühlborste nämlich zum dritten Mal berührt, wird die Produktion von Verdauungsenzymen gestartet, welche später durch Drüsen in den grünen Magen ausgeschieden werden. Besonders bemerkenswert ist, dass die Anzahl an Berührungen mit den Fühlborsten (vgl. Abb. 2) bestimmt, welche Menge an Verdauungsenzymen gebildet wird. Wenn ein großer Käfer aus dem Gefängnis zu entkommen versucht, berührt er die Fühlborsten öfter als eine Fruchtfliege. Die Pflanze erhält somit Information über die Größe der Beute und kann den Umfang des Enzymcocktails dementsprechend anpassen.
Doch nach drei Berührungen ist nicht Schluss! Werden die Fühlborsten insgesamt fünf Mal berührt, beginnt die Bildung von Natriumkanälen. Diese sind für die Aufnahme von Natrium in die Fangorgane zuständig. Das Natrium wird durch die Verdauungsenzyme aus der Beute gelöst. Auch hier konnte das Forschungsteam nachweisen, dass vermehrtes Berühren der Fühlborsten zu einem Anstieg in der Bildung der Natriumkanäle führt.
Es ist einerseits bemerkenswert, dass eine Falle die Fähigkeit hat, nach einer bestimmten Anzahl von Aktionspotentialen gewisse „Handlungen“ auszuführen. Andererseits fällt die geschickte Abstimmung der Enzyme und Natriumkanäle auf die Größe der Beute auf. Die Verarbeitung der Information aus den Fühlborsten erinnert an andere Regelkreisläufe der Natur, in denen komplexe Informationen verarbeitet werden und eine passende Reaktion darauf folgt. Es gilt abzuwarten, ob weitere Forschung zeigt, dass die Venusfliegenfalle in noch größeren Zahlenräumen operieren kann.
[Böhm J, Scherzer S et al. (2016) The venus flytrap Dionaea muscipula counts prey-induced action potentials to induce sodium uptake. Curr. Biol. 26, 286-295]
|
|
|
PDF-Version
Zur Entschlüsselung der geologischen Überlieferung werden gewöhnlich Prozesse herangezogen, die gegenwärtig auf der Erdoberfläche beobachtet werden. Diese Vorgehensweise – auch mit „die Gegenwart ist der Schlüssel zur Vergangenheit“ umschrieben – ist ein bedeutendes regulatives Prinzip* in der Erdgeschichtsforschung. Nyberg & Howell (2015) allerdings setzen ein Fragezeichen – bereits in der Überschrift ihres in Geology veröffentlichten Artikels: Is the present the key to the past? A global characterization of modern sedimentary basins. Ihre Analyse moderner Sedimentbecken erfolgte nicht im Rahmen dieser Fragestellung, vielmehr war es ihr Ergebnis, das zu einer Infragestellung des Prinzips führte.
Ziel der Studie von Nyberg & Howell war, Verteilung und Ausmaß bedeutender Sedimentbecken der kontinentalen Lithosphäre* zu kartieren, um eine repräsentative, globale Größe moderner Beckensysteme zu ermitteln und diese schließlich zur Gesteinsüberlieferung in Beziehung zu setzen.
Als moderne sedimentäre Becken definieren die Autoren Bereiche, die einer aktiven Subsidenz (Absenkung) unterliegen und in denen sich Sedimente ansammeln. Für die Kartierung und Quantifizierung haben nur Bereiche kontinentaler Lithosphäre mit quartären Sedimenten Berücksichtigung gefunden; Bereiche mit ozeanischer Lithosphäre (die meisten ozeanischen Becken) sowie unbekanntes Terrain (wesentliche Teile von Grönland und der Antarktis) sind ausgeschlossen worden. Zur Reproduzierbarkeit sind die Zielgebiete möglichst genau definiert worden: Einerseits Gebiete innerhalb der Platten mit Flächen > 10.000 km2 und niedrigem Relief (< 0,8°), z. B. Nord- und Ostsee; Schwarzes Meer, Kaspisches Meer, Aral-See und ihre umliegenden Gebiete; Teile Nordafrikas und Inneraustraliens (teilweise auch äolische Sedimente). Andererseits kleinräumigere Spezialgebiete (> 250 km2) wie Küsten- und intramontane Regionen, z.?B. Becken der Basin-und-Range-Provinz in den USA.
Im Ergebnis werden nur 16 % der Landoberfläche von Sedimentbecken eingenommen; die restlichen über 80 % sind Hochlandgebiete ohne zukünftiges geologisches Erhaltungspotential. Des Weiteren sind 60 % der modernen Beckenbereiche (der zuvor genannten 16 %) ariden Klimagebieten zuzurechnen, im Vergleich zu 27 % bezogen auf die gesamte Landoberfläche.
Das Ergebnis zeige an (S. 646), „dass der Anteil der Erdoberfläche, welcher innerhalb des terrestrischen Bereiches potentiell in der Gesteinsüberlieferung erhalten bleibt, nur 16 % betragen könne.“ Dies werfe „eine interessante Frage auf, [nämlich] wie repräsentativ die quantitative Verteilung moderner Analogien [hier Sedimentbecken, MK] für die Gesteinsüberlieferung (…) sind.“ Die Autoren schließen aus ihrer Analyse, dass die Kartierung und Quantifizierung moderner Becken die Unvollständigkeit der stratigraphischen Überlieferung herausstelle – wie [folglich] auch die Wichtigkeit eines vorsichtigen Umgangs mit der stratigraphischen Überlieferung, wenn eben angenommen wird, dass „die Gegenwart der Schlüssel zur Vergangenheit ist“ (S. 643).
Nyberg & Howell stellen mit ihrer konkreten Analyse die Anwendung dieses Prinzips bzw. seine Aussagekraft bei entsprechender Anwendung in Frage; es ist nicht ihr Anliegen, das (uniformitaristische) Prinzip wissenschaftstheoretisch zu diskutieren (vgl. Kotulla 2014, 2-21). Es ist anzumerken, dass die Autoren ihr (Gegenwarts-) Ergebnis auf die Vergangenheit übertragen – also im Grunde genommen das Prinzip anwenden. Aus ihren Ergebnissen hätte auch Folgendes geschlossen werden können: Die gegenwärtig auf der Erdoberfläche beobachtbaren Prozesse (mit Bezug auf Sedimentbecken und ihre Füllungen) sind mit denen der Vergangenheit nicht vergleichbar. Wo gibt es heute Sedimentationsräume, die beispielsweise denen des Karbons, Zechsteins oder Buntsandsteins vergleichbar wären? Streng genommen müssten die quartären Becken mit ihren Ablagerungen, die wesentliche Grundgröße der Analyse, separat betrachtet werden; denn auch sie sind – insbesondere die geologische Aktivität und Intensität betreffend – in ihrer Gesamtheit kein Abbild der Gegenwart.
Dass ihrem Schlusse zufolge die stratigraphische Überlieferung unvollständig ist bzw., mit Verweis auf Miall (2014), die Gesteinsüberlieferung eine unvollständige Überlieferung der geologischen Geschichte ist, auf diese (Erkenntnis-) Grenzen hat auch der Verfasser (Kotulla 2014, 2-01) aufmerksam gemacht.
|
Regulatives Prinzip: Regel oder Regelwerk zur Deutung der nicht beobachtbaren Vergangenheit, z. B. Uniformitarismus. Lithosphäre: Die Erdkruste sowie der obere, starre Teil des Erdmantels.
|
[Kotulla M (2014) Gültigkeit und Grenzen geologischer Zeitbestimmung. Online-Loseblattsammlung, Stand: 1. Ergänzungslieferung 03/2014, www.wort-und-wissen.de/loseblattsammlung.html • Miall A (2014) The emptiness of the stratigraphic record: A preliminary evaluation of missing time in the Mesaverde Group, Book Cliffs, Utah, U.S.A. Journal of Sedimentary Research 84, 457–469 • Nyberg B & Howell JA (2015) Is the present the key to the past? A global characterization of modern sedimentary basins. Geology 43, 643-646.]
|
|
|
PDF-Version
46 Autoren, allesamt Wissenschaftler der Tiefseebohr-Expedition 337, haben kürzlich gemeinsam in Science ihre Ergebnisse über die Erforschung der tiefen Biosphäre in Verbindung mit kohleführenden Sedimenten vorgestellt (Inagaki et al. 2015): Es existieren lebende Mikroorganismen mindestens bis in Tiefen von 2,5 km unter dem Meeresboden. Und: Taxonomische Gruppen in 2 km tiefen kohleführenden Sedimenten, die ursprünglich einen flachen küstennahen Ablagerungsraum besiedelten, scheinen seit ihrer Einbettung fortzubestehen – über einen Zeitraum von mehreren Zehner Millionen Jahren (Anm.: unter Zugrundelegung der geologischen Zeitskala, die mit radiometrischen Alterswerten geeicht ist).
Die 2012 mit dem derzeit modernsten Forschungsbohrschiff Chikyu im Rahmen des internationalen IOD-Programms (Integrated Ocean Drilling) durchgeführte Expedition 377 konnte einen neuen Tiefen-Rekord vermelden. An der Bohrlokalität Site C0020, auf dem westlichen pazifischen Kontinentalrand, 65 km östlich der Shimokita-Halbinsel (Japan), entstand in einer Wassertiefe von 1180 m das bislang tiefste Bohrloch einer wissenschaftlichen Tiefseebohrung – 2466 m (Expedition 337 Scientists 2013a).
 |
| Abb. 1: Kohlehaltiges Sediment. Bohrkern der IODP-Tiefseebohrung, Expedition 377, vom Kontinentalrand östlich Japan (Site C0020). (©Luc Riolon, MARUM/IODP) |
|
Die Bohrcrew reaktivierte ein bereits bestehendes Bohrloch (C9001) und setzte in einer Meeresbodentiefe von 647 m neu an. Wissenschaftlich beprobt wurden die rückgeführte Bohrspüle (Suspension, Bohrklein, Gase etc.) und die ab einer Tiefe von 1256 m punktuell gezogenen Kerne. Darüber hinaus wurden umfangreiche Bohrlochmessungen vorgenommen. Zur Identifizierung der Kontamination und ihrer quantitativen Berücksichtigung wurden spezifische Systeme und Verfahren entwickelt und eingesetzt.
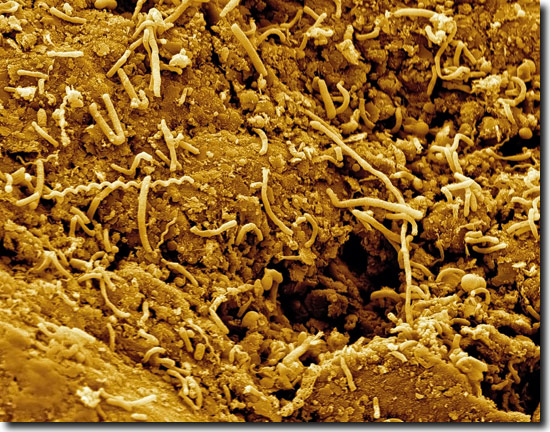 |
| Abb. 2: Kultur mit Mikrobenproben aus kohlehaltigem Sediment, das aus einer Meeresbodentiefe von 2 km stammt (vgl. Abb. 1). Im Bioreaktor wuchsen die Zellen auf dem der Kultur zugeführten Kohlenstaub und produzierten Methan. Elektronenmikroskop-Aufnahme, etwa 5.000-fache Vergrößerung. Bildbreite: ca 25 mm. (© Hiro Imachi, JAMSTEC) |
|
Die durchbohrten Ablagerungen bestehen hauptsächlich aus unterschiedlich verfestigten Sand-, Silt- und Tonsteinen sowie Konglomeraten; sie wurden in vier (lithologische) Einheiten gegliedert. Im tieferen Bereich herrschen Temperaturen von ~ 40 ° bis 60 °C. Der Tiefenabschnitt von 1826-2046 m (Einheit III) zeichnet sich durch Einschaltungen von Kohle- bzw. kohlehaltigen Schichten aus (Abb. 1), darunter zwölf bedeutendere, zehn mit Dicken von 30 bis 140 cm und zwei mit 350 bzw. 730 cm (Expedition 337 Scientists 2013b, Tab. T10). Die biostratigraphische Einordnung mittels Diatomeen (Kalkalgen, nur am Top) und Palynomorphen (Sporen und Pollen) ist recht grob: Am Top (647 m) oberes Pliozän und an der Basis (2466 m) oberes Oligozän/unteres Miozän; Einheit III oberes Oligozän/unteres bis mittleres Miozän. Der Sedimentationsraum wird in seiner kontinuierlichen Entwicklung als zunächst küstennah und stark wechselnd (terrestrisch, intertidal, flachmarin) gedeutet, sodann als marin und zunehmend küstenfern (heute Kontinentalrand). Demzufolge entstanden die Kohleschichten in 2 km Tiefe, „(…) als große Mengen organischen Materials, das aus Landpflanzen stammt, in Küstennähe abgelagert wurden“ (MARUM) und das Sedimentbecken insgesamt absank. Beckenabsenkung (Subsidenz) und Sediment-Input müssen weitestgehend in Balance gestanden haben (Expedition 337 Scientists 2013a, 13).
Die Forscher entdeckten über die gesamte Tiefe intakte mikrobielle Zellen. Die Zellkonzentration nahm erwartungsgemäß mit der Tiefe ab, im „flacheren“ Meeresbodenuntergrund (bis 365 m) von ~109 zu ~107 Zellen pro cm3. Im „tieferen“ Meeresbodenuntergrund (> 1,5 km) lag sie mit ~ 102 bis 103 Zellen pro cm3 aber deutlich unter der Erwartung. Auffällig dagegen waren lokal höhere Konzentrationen in den kohleführenden Sedimenten.
Des Weiteren kann sogar von einer mikrobakteriellen Aktivität im gesamten Untergrund bis 2,5 km Tiefe ausgegangen werden. Das zeigten spezifische und kontinuierliche Methananalysen an, die auf eine Methanerzeugung durch Mikroben hinweisen. Die Autoren mutmaßen, dass wahrscheinlich der größte Teil des Methans auf diese Weise entstanden sein könnte. Insbesondere wirke das kohlige Substrat (erhöhte Zellkonzentrationen) stimulierend auf Mikrobenwachstum und Methanproduktion. Den indirekten Nachweis lieferte ein Laborexperiment: In einem Bioreaktor waren Proben der methanbildenden Mikroben-Vergesellschaftungen aus den kohleführenden Sedimenten bei 40 °C mit Kohlestaub, der Hauptenergiequelle, angereichert worden. Die Bakterien wuchsen und produzierten durch Umwandlung von Kohle Methan (Abb. 2). Die in der Kultur gewachsenen Methanogene seien mit Methanobacterium subterraneum und M. formicicum eng verwandt, Arten, die kürzlich in terrestrischen Kohleschichten entdeckt worden waren.
Gensequentielle Analysen (auf Basis des bakteriellen Gens 16S rRNA) zeigten deutliche Unterschiede in der Zusammensetzung der Mikrobengemeinschaften im „flacheren“ und „tieferen“ Meeresbodenuntergrund. So würden die Gemeinschaften der „Tiefe“ von Bakteriengruppen dominiert, die von terrestrischen Böden bekannt sind; insbesondere würden viele operationale taxonomische Einheiten (OTU) eine hohe Sequenz-Übereinstimmung mit denen von Waldböden aufzeigen.
Dieses Milieu im tiefen Meeresbodenuntergrund scheint, so schließen Inagaki et al. (2015, 424), „einige der taxonomischen Gruppen, die ursprünglich den flachen Ablagerungsraum besiedelten, erhalten haben und seitdem zum Kohlenstoffkreislauf beigetragen haben“. Die Zeitdauer dieses vermuteten Fortbestandes – rund „20 Millionen Jahre“ (MARUM) – wird allerdings weder diskutiert noch problematisiert.
[Expedition 337 Scientists (2013a) Expedition 337 summary. In: Inagaki F, Hinrichs K-U, Kubo Y (eds) Proc. IODP 337, Tokyo (Integrated Ocean Drilling Program Management International, Inc.). doi:10.2204/iodp.proc.337.101.2013 • Expedition 337 Scientists (2013b) Site C0020. In: Inagaki F, Hinrichs K-U, Kubo Y (eds) Proc. IODP 337, Tokyo (Integrated Ocean Drilling Program Management International, Inc.). doi:10.2204/iodp.proc.337.103.2013 • Inagaki F, Hinrichs K-U & weitere 44 Autoren (2015) Exploring deep microbial life in coal-bearing sediment down to ~2.5 km below the ocean floor. Science 349, 420-424 • MARUM (2015) Auf der Suche nach den Grenzen des Lebens. Pressemitteilung des Zentrums für Marine Umweltwissenschaften, Universität Bremen, vom 24. Juli 2015]
|
|
|
Anfang der 1990er-Jahre hat Schopf (1993) mikroskopische Strukturen in Feuersteinen (Apex Chert) aus Australien beschrieben. Diese Gesteine werden zu den ältesten Gesteinen gezählt, die auf der Erde vorkommen. Ihr Alter wird mit 3,46 Milliarden radiometrischen Jahren angegeben. Seit der ersten Beschreibung werden die Interpretationen der Befunde kontrovers diskutiert, d. h. es gibt Autoren, die ausschließlich physikalisch-chemische Ursachen für die Entstehung dieser Strukturen anführen und sie als Pseudofossilien (Brasier et al. 2002) ansprechen, während andere Wissenschaftler Befunde und Argumente anführen, die für einen biologischen Ursprung dieser Mikrofossilien sprechen.
Jetzt haben Bower et al. (2016) ursprüngliche Proben, die im Natural History Museum in London archiviert sind, einer umfangreichen Analyse unterzogen. Dabei haben sie neu entwickelte Methoden eingesetzt, in welchen Signale von Raman-Spektren – genauer das Verhältnis aus zwei Signalen – zur Erzeugung von hochaufgelösten Bildern genutzt werden. Diese Darstellungen ermöglichen die Feststellung der Orientierung von Quarzkristallen in den untersuchten Proben.
Diese Methode wurde, nachdem sie vorher an bekannten Proben evaluiert worden war, auf eines der Fossilien angewendet, die Schopf als Holotypus für die Art Eoleptonema apex beschrieben hatte, also eines, das maßgeblich für die Beschreibung dieser Art ist. Wird nur ein Raman-Signal (461 cm-1) für die bildliche Darstellung der Verteilung von Quarz benutzt, so zeigt sich eine ziemlich gleichförmige Verteilung bis auf die unmittelbare Umgebung der als Mikrofossil beschriebenen Struktur und innerhalb derselben. Die Bilder auf Basis von für Kohlenstoff spezifischen Signalen zeigen, dass dieses Element in und neben der fraglichen Struktur zu finden ist. Die Abbildungen, die mit der von den Autoren entwickelten Methode erstellt wurden, legen nahe, dass einzelne Quarzkristalle von der als Mikrofossil beschriebenen Struktur durchschnitten wurden.
Die von Bower et al. vorgelegten Daten werden von den Autoren so interpretiert, dass das von Schopf als E. apex beschriebene Mikrofossil sehr viel wahrscheinlicher einen Riss darstellt, der entstand, während der Kristall aus einer Lösung gewachsen ist, die reich an Silikat und Kohlenstoff war. Der spektrometrisch nachgewiesene Kohlenstoff entspricht dann einer Rissfüllung.
Über den Ursprung des Kohlenstoffs machen die Autoren keine verbindlichen Aussagen. Dieser könnte in der Tat biologischen Ursprungs, aber auch unter hydrothermalen Bedingungen anorganisch entstanden sein.
Bowner et al. (2016) plädieren dafür, dass vor einer sorgfältigen und umfassenden Analyse der mikroskopischen Strukturen in Feuerstein diese als Pseudofossilien aufgefasst werden sollten, also als Strukturen, die nur so aussehen, als wären sie Mikrofossilien und damit Hinweise auf Lebewesen. Damit ist an einem Beispiel der von Schopf in einer aufsehenerregenden Veröffentlichung beschriebenen Mikrofossilien aus den ältesten verfügbaren Gesteinen gezeigt, dass deren Natur nach wie vor ungeklärt ist. Das bedeutet, dass man bei der Interpretation gerade sehr alter Strukturen sehr vorsichtig vorgehen sollte.
[Bower DM, Steele A, Fries MD, Green OR & Lindsay JF (2016) Raman imaging spectroscopy of a putative microfossil from the ~3,46 Ga Apex chert: insights from quartz grain orientation. Astrobiol. 16, 169-180 • Brasier MD, Green OR, Jephcoat AP, Van Kranendon MJ, Lindsay JF, Steele & Grassineau NV (2002) Questioning the evidence for the earth‘s oldest fossils. Nature 416, 76-81 • Schopf JW (1993) Microfossils of the Early Archean Apex chert: new evidence of the antiquity of life. Science 260, 640-646.]
|
|
 |
| Abb. 1: Zirkone zählen zu den ältesten Mineralien in der Erdkruste. Sie bestehen hauptsächlich aus Zirkoniumsilikat und können größere Mengen an Verunreinigungen enthalten. Das hier abgebidete Exemplar wurde von Forschern der Senckenberg Gesellschaft für Naturforschung im permischen „Versteinerten Wald“ von Chemnitz aus einer Aschelage geborgen und zur radiometrischen Datierung genutzt. (© Senckenberg) |
|
Hinweise auf Leben kann man in Gesteinen in verschiedener Form weit in die Vergangenheit zurückverfolgen. In Fossilien finden wir oft deutliche Spuren auf vergangene Organismen; mikroskopische Strukturen in Gesteinen, sogenannte Mikrofossilien, lassen sich noch weiter zurück verfolgen, sind aber auch schwerer zu interpretieren, und die Fachleute diskutieren sie kontrovers. Chemische Besonderheiten, die als Hinweise auf Leben verstanden werden, finden sich in den ältesten Gesteinen, die man bisher auf der Erde findet; aber auch in diesem Fall ist der Interpretationsspielraum sehr groß.
Im Westen Australiens (Jack Hills, Narryer-Gneis Terran) werden sehr alte Gesteinsformationen (Metasedimente) beschrieben, in denen Zirkonkristalle eingeschlossen sind, die ursprünglich aus noch älteren Gesteinen stammen (detritisch). Einigen dieser Zirkone wird aufgrund radiometrischer Bestimmungen ein Alter von mehr als 4 Milliarden Jahren zugesprochen. Bell et al. (2015) haben 656 Zirkonkristalle mit einem radiometrischen Alter von mehr als 3,8 Milliarden Jahren auf Kohlenstoff (C)-haltige Einschlüsse untersucht. Dabei fanden sie einen Zirkon, dem sie aufgrund von Uran(U)-Blei(Pb)-Messungen ein U-Pb Alter von 4,10 +/–0,01 Milliarden Jahren zuweisen. Dieser Kristall enthält in einem ungestörten Bereich (keine Risse) graphitartige C-Einschlüsse, von welchen der Gehalt an stabilen C-Isotopen C-12 und C-13 bestimmt wurde. Daraus lässt sich ein δ13C-Wert bestimmen, der im Vergleich zu einem Standard (Pee Dee Belemniten Standard: δ13CPDB) die Anreicherung bzw. Verarmung an C-13 angibt. Ein im Vergleich zum Standard deutlich verringerter 13C-Gehalt wird aufgrund bisheriger Erfahrung als Hinweis auf eine biologische Quelle des Kohlenstoffs interpretiert; dabei wird die enzymatische Aktivität in lebenden Zellen für den Effekt verantwortlich gemacht. So wurden Partikel mit Proben aus Grönland mit einem radiometrischen Alter < 3,7 Milliarden Jahren aufgrund von δ13C-Werten als frühe Hinweise auf Leben interpretiert (Mojzsis et al. 1996, Rosing 1999).
Bell et al. (2015) ermittelten einen δ13C-Wert von -24 +/- 5 ‰, der damit in einem Bereich liegt, der für eine biologische C-Quelle spricht. Die Autoren diskutieren umfänglich alternative anorganische Erklärungen für die Verarmung an 13C und verteidigen dabei die Plausibilität eines biologischen Ursprungs des Kohlenstoffs in der von ihnen präsentierten Probe.
Angesichts des Interpretationsspielraums der Daten und der Tatsache, dass nur ein einziger Zirkonkristall diese Art des Einschlusses aufweist, ist die Aussagekraft der Arbeit von Bell et al. (2015) begrenzt und bedarf einer Bestätigung durch weitere zukünftige Untersuchungen. Dennoch liefert sie möglicherweise den derzeit ältesten Hinweis auf Leben mit > 4,1 • 109 radiometrischen Jahren rund 300 • 106 Jahre älter als bisher bekannt. Damit weisen die Autoren wieder einmal darauf hin, dass Leben auf der Erde sehr früh nachweisbar zu sein scheint, was die Erklärung seiner möglichen natürlichen Entstehung auf der Erde vor eine große Herausforderung stellt.
[Bell EA, Boehnke P, Harrison TM & Mao WL (2015) Potentially biogenic carbon preserved in 4.1 billion-year-old zirkon. Proc. Natl. Acad. Sci. 112, 14518–14521 • Mojzsis SJ, Arrhenius G, McKeegan KD, Harrison TM, Nutman AP & Friend CRL (1996) Evidence of life on earth by 3800 million years ago. Nature 384, 55-59 • Rosing MT (1999) δ13C-depleted carbon microparticles in > 3700-Ma sea floor sedimentary rocks from West greenland. Science 283, 674-676]
|
|
|
 |
