 |
Fossiler Fund eines Säugetiers mit prächtigem Haarkleid
Konvergente Entstehung eines weiteren Schlüsselmerkmals
von Henrik Ullrich
Studium Integrale Journal
23. Jahrgang / Heft 1 - Mai 2016
Seite 34 - 38
|
 |
Zusammenfassung: Der Fossilfund eines eutriconodonten Säugetiers aus den unteren Kreideschichten Spaniens dokumentiert ein evolutionstheoretisches Dilemma. Schlüsselmerkmale der Säugetiere sind bereits bei Individuen aus den am ältesten datierten Fundhorizonten nachzuweisen. Andererseits zwingt die Merkmalsverteilung bei fossilen und rezenten Säugetieren zu dem Schluss, dass Schlüsselmerkmale mehrfach unabhängig voneinander mit erstaunlicher morphologischer Identität entstanden sind. Eine kausale evolutionstheoretische Erklärung für diese Befunde fehlt.
| |
|
Schlüsselmerkmale der Säugetiere: Haut und Haare
Säugetiere unterscheiden sich von Reptilien, die als ihre evolutionäre Ahnen gelten, durch den Besitz von besonderen Merkmalen. Dazu zählen die Mittelohrknochen (Hammer, Amboss, Steigbügel), Milchdrüsen, Fell bzw. Haare, die Warmblütigkeit oder speziell bei den Plazentatieren das Reifen und Heranwachsen der Nachkommen in der Gebärmutter mit der Ausbildung einer Plazenta. Über viele Jahrzehnte waren Evolutionsbiologen überzeugt, dass diese Schlüsselmerkmale im Verlauf der Evolution, also beim Übergang von Reptilien zu den ersten Säugetierformen, nur einmal (monophyletisch) entstanden und von allen folgenden Säugetierformen übernommen wurden.
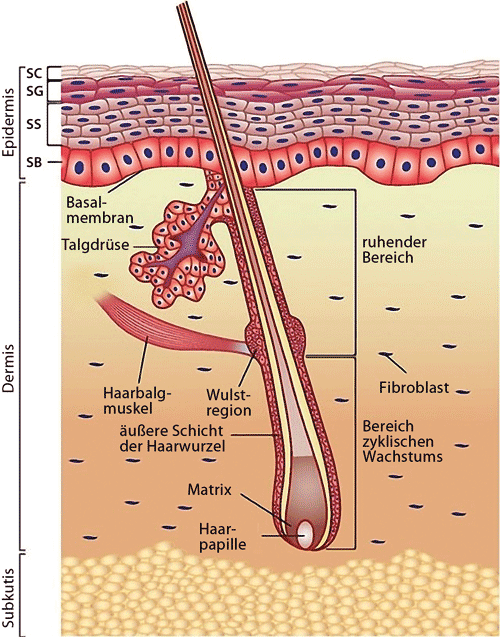 |
| Abb. 1: Schematische Darstellung des Aufbaus der Haut des Menschen. SB stratum basale = Basalzellschicht, SC stratum corneum, SG stratum granulosum, SS stratum spinosum. Erläuterungen siehe Text. (Wikimedia Commons, Cc-by-3.0) |
|
Die Haut (Integument) der Säugetiere stellt einen Merkmalskomplex dar, welcher die funktionelle und anatomische Basis für die Existenz von Schlüsselmerkmalen bereitstellt (z.?B. für die Warmblütigkeit, für das Fell bzw. die Haare). Mit ihren Anhangsorganen (z. B. Haare, Nägel, Stacheln, Hautschilde, Drüsen, Brustdrüse) ist die Haut das größte Organ der Säugetiere. Beim erwachsenen Menschen bedeckt sie eine Oberfläche von 1,5-2,0 Quadratmetern und erreicht ein Gewicht von durchschnittlich 14 Kilogramm (ohne Unterhaut).
Charakteristisch für die Haut (Abb. 1) ist ihr schichtförmiger Aufbau aus Oberhaut (Epidermis) und der Lederhaut (Dermis oder Corium). Der Haut unterlagert findet sich die Unterhaut (Subcutis). Die Epidermis, welche ontogenetisch aus dem äußeren Keimblatt, dem Ektoderm, hervorgeht, zeigt große regionale Unterschiede auf makroskopischem und mikroskopischem Niveau innerhalb eines Individuums (z. B. Handfläche ohne Drüsen und Behaarung, Kopfhaut mit Haaren und Drüsen).
Die Lederhaut entsteht ontogenetisch aus dem mittleren Keimblatt, dem Mesenchym, und steuert maßgeblich die Differenzierung, Umformung und Proliferation (Vermehrung der jeweiligen Zelltypen) der Epidermis und der Hautanhangsstrukturen (z.?B. der Haare). Die Epidermis, die sich aus dem äußeren Keimblatt, dem Ektoderm, ontogenetisch ableitet, grenzt sich durch eine Basalmembran von der Lederhaut ab. Diese Epithelschicht stellt einen zentralen Ort der epithelio-mesenchymalen Wechselwirkungen dar, die ihrerseits wesentlich zur Bildung der o. g. spezialisierten Strukturen der Haut beitragen.
 |
| Abb. 2: Innerartliche Variabilität der Haartypen und Haarformen am Beispiel einer lokalen Haarsammlung von Einwohnern aus dem Ort Sulzbach. (Wikimedia Commons) |
|
In die Hautschichten wandern während der Ontogenese Zellen aus anderen Körperregionen ein und beeinflussen ebenfalls die Entwicklung und Differenzierung der Haut und ihrer Anhangsgebilde maßgeblich mit. Beispielsweise entstammen in die Haut einwandernde Neuralleistenzellen (neural crest cells) aus der Umgebung der embryonalen Anlage des zentralen Nervensystems. Aus diesen gehen unter anderem Pigmentzellen (Melanozyten) hervor, die für die Färbung der Haut und der Haare verantwortlich sind. Ein anderes Beispiel sind die dem Knochenmark entstammenden Langerhanszellen, die im Rahmen der Immunabwehr (T-Zellantwort) der Haut eine wichtige Funktion innehaben.
Unter dem induzierenden und steuernden Einfluss der Dermis entwickeln sich wie bereits erwähnt die für Säugetiere typischen Haare, Stacheln, Nägel und Hautschilde. In der Epidermis vorhandene spezialisierte Zellen, die Keratinozyten, produzieren dafür ein besonderes Protein, das Keratin, das den Zellen ihre strukturelle Integrität und später ihre morphologische Charakteristik vermittelt. Die Keratinisierung basiert auf einer komplexen, genetisch kontrollierten und programmierten Serie von morphogenetischen (formbildenden) Veränderungen, die ihren Motor im Bereich der Kontaktzone zwischen Epidermis und Dermis (z. B. Basalmembran) hat (vgl. O’Rahilly & Müller 1999). Neueste Untersuchungen haben darüber hinaus gezeigt, dass mehr als 50 verschiedene Varianten des Keratinproteins beim Säugetier existieren und diese in engsten Wechselwirkungen mit Mitochondrien – den Kraftwerken der Zellen – stehen, um den differenzierten funktionellen Ansprüchen gerecht zu werden (Kumar et al. 2015).
Die ersten Haare entstehen beim Menschen am Ende der Embryonalperiode in den Augenbrauen, der Oberlippe und dem Kinn. Zu Beginn sind Einstülpungen der Epidermis (Follikel) zu sehen, die Stellen künftiger Haare und Talgdrüsen; an der tiefsten Stelle dieser Einstülpung entsteht der Haarbulbus. Durch dessen Kontakt mit dem dermalen Mesenchym werden zentrale Anteile der Follikel keratinisiert, der Haarschaft dringt durch das basale Zellwachstum im Haarbulbus zur Oberfläche vor. Die Färbung der Haare erfolgt durch Melanozyten aus der Neuralleiste (s. o.). Die ersten Haare (Lanugobehaarung) werden zur Zeit der Geburt abgestoßen und durch die Terminalhaare ersetzt.
Ab einer Haardichte von 50 bis 400 Haaren je Quadratzentimeter spricht man von einem Fell, eine noch stärkere Behaarung nennt man Pelz. Das Fell besteht in der Regel aus zwei Anteilen, dem Deckhaar (Oberhaar) und dem Wollhaar. Stacheln wie beim Igel entstehen ontogenetisch etwas später als die Haare durch die Fusion mehrerer keratinisierter Tubuli, die ein starres hohles Haar formen. Eine besondere Spezialisierung von Haaren bei Säugetieren sind die hochsensiblen Tasthaare (Vibrissen).
Ein auffälliges Merkmal heutiger Säugetiere stellt der außergewöhnliche Polymorphismus (Vielgestaltigkeit) der Haare dar (Abb. 2). Diese morphologischen Varianten finden sich schon bei Individuen, die systematisch eng zueinander gehören, darüber hinaus auch bei einem einzelnen Individuum an unterschiedlichen Körperregionen und während der individuellen Ontogenese in aufeinanderfolgenden Entwicklungsstadien.
|
|
Das Fossil Spinolestes xenarthrosus
Martin et al. (2015) beschreiben das Fossil eines außergewöhnlich gut erhaltenen Säugetiers aus der ausgestorbenen Gruppe der Eutriconodonten, das sie in Schichten der Unterkreide in Spanien (Las Hoyas) fanden, auf der Basis von Leitfossilien der Barremium-Stufe zuordneten und auf 125-127 Millionen radiometrische Jahre (MrJ) vor heute datierten. Es fand sich ein komplettes Skelett mit Gelenken, einer außergewöhnlichen Erhaltung von Spuren der Haut, der Haare, keratinhaltigen Hautstacheln und Spuren von Organen des Brustkorbes und des Bauchraumes. Neben anderen Merkmalen fanden sich ein ossifizierter Meckelknorpel (vgl. Ullrich 2012) und spezialisierte Gelenkverbindungen wie bei heutigen Nebengelenktieren im Bereich der unteren Wirbelkörper, welche die Wirbelsäule bei Bewegung stabilisieren. Der Name Spinolestes xenarthrosus verbindet diese anatomischen Besonderheiten („Spino“: Stachel, „Xenarthrosus“: Nebengelenk).
Wir wollen uns an dieser Stelle auf die Besonderheiten des Haarkleides des fossilen Fundes konzentrieren. Abdrücke von fossilem Fell, also einer kompletten Bedeckung der Haut mit Haaren, sind aus dem mittleren Jura von einem im Wasser lebenden Ursäuger bekannt (Castorocauda lutrasimilis, datiert auf 162 Millionen radiometrische Jahre, Ji et al. 2006). Fossilisierte Haare sind in Schichten des untersten Tertiärs (ca. 60 MrJ, Paläozän) gefunden worden (Meng et al. 1997). Der vorliegende Fund mit einem angegebenen Alter von 125-127 MrJ präsentiert zahlreiche fossile Haarstrukturen mit unterschiedlichen Entwicklungsgraden, die auf makroskopischer und mikroskopischer Ebene den Haarstrukturen bei heutigen Säugetieren exakt entsprechen. Diese Entsprechung gilt, so Martin et al., für den individuellen Polymorphismus der Haare und auch für die zugrundeliegenden ontogenetischen Prozesse und die entwicklungsgeschichtlichen Stadien der Haarentwicklung.
Zum ersten Mal werden hier zylindrische Strukturen beschrieben (Protostacheln –„Protospines“), die mikroskopisch analog den Mikrostacheln bei heutigen Stachelmäusen sind. Sie befanden sich bei Spinolestes xenarthrosus im Rückenbereich über den Dornfortsätzen der Wirbelkörper.
 |
| Abb. 3: Rekonstruktion des von Martin et al. beschriebenen Fossil Spinolestes xenarthrosus. Spinolestes war ca. 20 cm lang und erreichte eine Schulterhöhe von ca. 5,5 cm. Weitere Erläuterungen siehe Text. (Aus Martin et al. 2015) |
|
Die exquisite fossile Erhaltung von vielen Hautstrukturen zeigt deutlich eine regionale Differenzierung der Haare und der Mähne, es finden sich Deckhaare, 3-5 mm lang (in der seitlichen Körperregion, am Hals und am Schulterblatt) und lange zarte Haare im Rückenbereich und am Schwanz. Die übrigen Körperabschnitte wurden von einem dichten Unterfell bedeckt.
 |
| Der vorliegende Fund präsentiert zahlreiche fossile Haarstrukturen mit unterschiedlichem Entwicklungsgraden, die den Haarstrukturen bei heutigen Säugetieren exakt entsprechen. |
 |
|
Eindrucksvoll sind auch folgende mikroskopische Details: Es gelang den Forschern, am Fossil primäre und sekundäre Haarstrukturen mit Follikeln zu identifizieren, wie sie in allen drei modernen Säugertiergruppen vorkommen. Z. T. ließen sich bei Spinolestes xenarthrosus auch Spuren von durch Pilzbefall veränderten Haaren wie bei heutigen Säugern dokumentieren.
„Die Haut dieses neuen mesozoischen Säugetiers vereinigt die gleiche Kombination von Hautanhangsorganen wie jene, die unabhängig in der gleichen Epoche in anderen Gruppen der Kronensäugetiere evolutionär entstanden sind mit einer ähnlich großen strukturellen Variabilität wie bei den heutigen Säugetieren“ (Martin et al. 2015, 380).
|
Homologie: Gleichwertigkeit (z. B. gleiches Bauprinzip) ähnlicher, z. T. aber auch unähnlicher Strukturen, Organe oder Prozesse; in der Evolutionsbiologie als Belege für gemeinsame Abstammung interpretiert. Homoplasien: Bauplanähnliche Strukturen, die trotz ihrer Ähnlichkeit als mehrfach unabhängig entstanden interpretiert werden müssen. Dazu gehören Konvergenzen, Parallelentwicklungen und Rückentwicklungen. Konvergenz: gleichförmige Entwicklung ähnlicher Strukturen und Funktionen einzelner Organe nicht abstammungsmäßig verwandter Organismen.
|
|
|
Konsequenzen für die evolutions-theoretische Modellbildung der Säugetierentstehung
Für die Autoren steht außer Frage: In einem evolutionären Kontext muss davon ausgegangen werden, dass die Haut mit ihren Anhangsorganen (z. B. Haare, Stacheln) bereits sehr früh von den ersten Ursäugetierformen evolutionär erworben wurde. Schon im Mesozoikum waren nicht nur unspezialisierte Haare vorhanden, sondern das gesamte Spektrum ihres Polymorphismus und der heute beobachtbaren individuellen ontogenetischen Entwicklungsschritte.
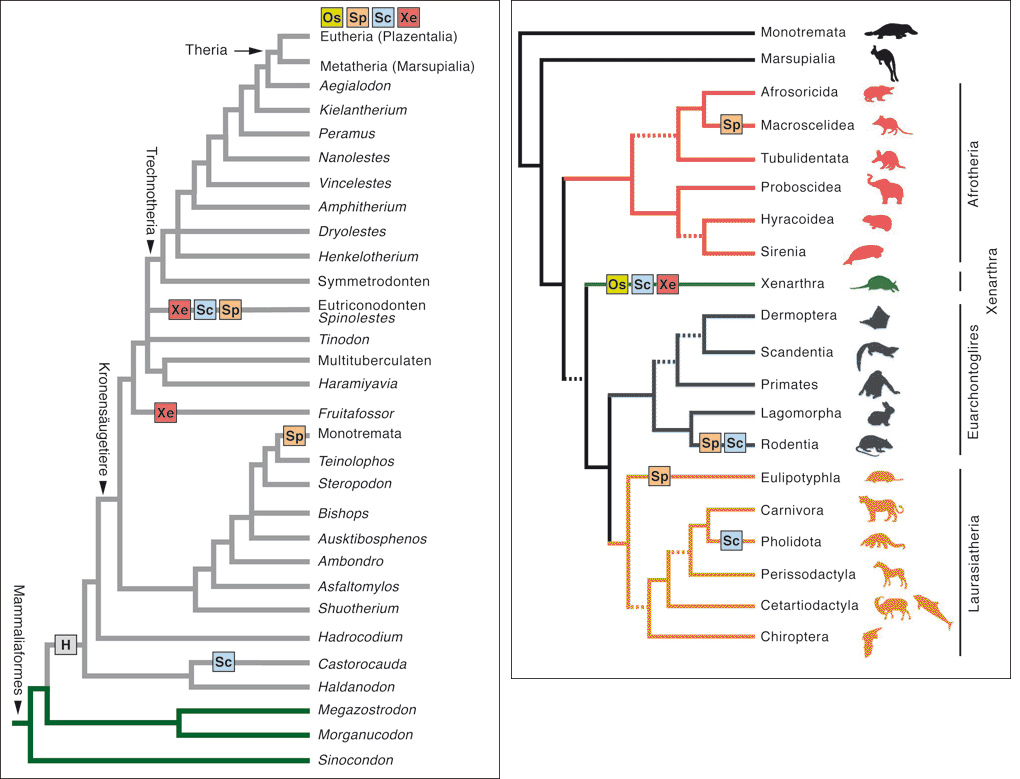 |
Abb. 4: Konvergenz des Auftretens von spezifischen Hautanhangsstrukturen bei Säugetieren. Systematik fossiler (links) und rezenter (rechts) Säugetiere unter Berücksichtigung fossiler Nachweise des ersten konvergenten Auftretens von Haaren und Hautanhangsstrukturen sowie des konvergenten Erscheinens der für die Nebengelenktiere typischen Wirbelkörperformen. Die Position des in diesem Artikel beschriebenen fossilen Fundes Spinolestes (Eutrinoconodont) ist besonders markiert.
H Erster Nachweis von Haaren und Sporen/Stacheln, Sc keratinisierte / verhornte Schilder bzw. Platten, Sp Hautstacheln, Os ossifizierte Hautschilder, -platten, Xe Für Nebengelenktiere (Xenarthra) typische Konfiguration der hinteren Wirbelkörper. (Nach Martin et al. 2015, Luo 2007, Springer et al. 2004) |
|
Demgegenüber dokumentiert der Fossilbefund bislang nicht, über welche anatomischen Zwischenschritte die Haut der Säugetiere sich mit ihren Anhangsorganen aus der Haut reptilienartiger Vorgänger entwickelt haben könnte. Ein kennzeichnendes Merkmal der rezenten Reptilien ist ihre trockene, schleimlose und drüsenarme Haut, die von Schuppen, Hornplatten oder Panzer bedeckt wird. Diese äußere Schicht wird bei Schlangen und Echsen regelmäßig durch Häutung erneuert.
|
| Neben anderen Schlüsselmerkmalen der Säugetiere müssen auch die Anhangsorgane der Haut mehrfach konvergent entstanden sein. |
|
|
Als weitere Schlussfolgerung aus dem beschriebenen Befund und aus dem Vergleich mit weiteren fossilen Säugetierformen folgt für Martin et al. die Tatsache, dass neben anderen Schlüsselmerkmalen der Säugetiere (z.?B. Mittelohrknochen) auch die Anhangsorgane der Haut mehrfach konvergent* innerhalb der einzelnen Säugetierlinien evolutionär entstanden sind. Und dies mit identischen Ergebnissen auf makroskopischer, mikroskopischer und ontogenetischer Ebene. Diese Aussage ist nicht trivial oder nebensächlich. Die Konvergenz betrifft nicht lediglich ein Einzelmerkmal, sondern ontogenetisch, funktionell und anatomisch hoch vernetzte Merkmalskomplexe, auf denen fundamentale Eigenschaften der Säugetiere basieren (s. o.).
2007 haben bereits Luo et al. auf die sich immer mehr abzeichnende und unerwartete Befundkonstellation im Hinblick auf die Evolution der Schlüsselmerkmale der Säugetiere hingewiesen und von einem neuem Paradigma evolutionärer Theorienbildung gesprochen: „Das aufstrebende neue Paradigma besagt, dass durch aufeinanderfolgende Diversifikationen innerhalb der mesozoischen Säugetiere es vielen ausgestorbenen Linien möglich wurde, unterschiedliche ökologische Lebensräume zu erobern – genauso wie in der känozoischen Säugetierdiversifikation (aber nicht so erfolgreich). Diese erfolgten in unabhängigen evolutionären Experimenten, basierend auf tiefgreifenden Homoplasien* der ontogenetischen Entwicklungen und konvergenten funktionellen und ökologischen Anpassungen“ (Luo et al. 2007, 1017, 1018; Hervorhebung hinzugefügt).
|
|
Diskussion
Das Mesozoikum (252-66 MrJ) gilt immer noch als das Zeitalter der Dinosaurier, obwohl die dieser Spanne zugeordneten geologischen Schichtfolgen auch eine bemerkenswerte ökomorphologische Vielfalt ausgestorbener, hochspezialisierter Säugetierformen bergen. Im Szenario der Evolution entstanden die Säugetiere mit ihren charakteristischen Schlüsselmerkmalen zeitgleich mit den Dinosauriern im Mesozoikum. Über viele Jahrzehnte im 20. Jahrhundert galten diese hochkomplexen Schlüsselmerkmale für die Fachwelt als einmalige (monophyletische) evolutionäre Erfindungen, die eine monophyletische evolutionäre Abstammungsfolge der Säugetiere im Rahmen der Synthetischen Evolutionstheorie (SET) begründeten.
|
| Die für die Konvergenzen verantwortlichen Mechanismen sind auf molekulargenetischer Ebene völlig unklar. |
|
|
In den letzten zwei Jahrzehnten mussten auf der Grundlage zahlreicher Fossilfunde ausgestorbener Säugetierformen aus dem Mesozoikum die klassischen phylogenetischen Konzeptionen der SET aufgegeben und eine komplett neue Systematik der Säugetiere entworfen werden (Ullrich 2013). Der Fossilbefund – unter der Leitidee „Evolution“ interpretiert – lässt gegenwärtig nur den Schluss zu, dass mehrfach und unabhängig voneinander Säugetierformen evolutionär entstanden, die nahezu identische und konvergente Merkmalskomplexe aufweisen (siehe Glossar), die ihnen eine optimale Anpassung in verschiedenen Ökosystemen (im Wasser, auf Bäumen oder als flugfähige Individuen) ermöglichten (sog. identische Ökomorphotypen). Die Annahme der konvergenten Entstehung dieser Merkmalskomplexe der Säugetiere schließt auch die komplexen Schlüsselmerkmale der Säugetiere ein. Während diese Konvergenzen bei Säugetieren unter den Evolutionsbiologen nicht mehr zur Debatte stehen, sind die dafür verantwortlichen Mechanismen auf molekulargenetischer Ebene völlig unklar. Es sind nicht einmal ansatzweise Mechanismen bekannt, die in der Lage wären, über Jahrmillionen immer wieder ungezielt die „tiefgreifenden Homoplasien der ontogenetischen Entwicklungen und konvergenten funktionellen und ökologischen Anpassungen“ (Luo et al. 2007) hervorzubringen. Zum anderen fehlt es regelmäßig an fossilen Belegen für die postulierten mehrfachen konvergenten Entwicklungswege, aus denen u. a. die Schlüsselmerkmale der Säugetiere hervorgegangen sein sollen.
Der aktuelle Fund eines eutriconodonten Säugetiers aus den unteren Kreideschichten Spaniens dokumentiert dieses evolutionstheoretische Dilemma eindrücklich am Beispiel außergewöhnlich gut überlieferter fossiler Spuren eines wichtigen Schlüsselmerkmales der Säugetiere: der Haut und ihrer Anhangsorgane.
|
Literatur
- Ji Q et al. (2006)
- A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals. Science 311, 1123-1127.
- Luo ZX et al. (2007)
- A new eutriconodont mammal and evolutionary development in early mammals. Nature 446, 288-293.
- Kumar V et al. (2015)
- A keratin scaffold regulates epidermal barrier formation, mitochondrial lipid composition, and activity. JCB 211, 1057-1075.
- Martin T et al. (2015)
- A Cretaceous eutriconodont and integument evolution in early mammals. Nature 526, 380-384 .
- Meng J & Wyss AR (1997)
- Multituberculate and other mammal hair recovered from Palaeogene excreta. Nature 385, 712-714.
- O’Rahilly R & Müller F (1999)
- Human Embryology and Teratology. New York.
- Springer MS et al. (2004)
- Molecules consolidate the placental mammal tree. Trends Ecol. Evol. 19, 430-438.
- Thanukos A (2008)
- Bringing homologies into focus. Evo. Edu. Outreach 1, 498-504.
- Ullrich H (2012)
- Verwirrende Konvergenzen. Zur Evolution des Mittelohres der Säugetiere. Stud. Integr. J. 19, 20-33.
- Ullrich H (2013)
- Moleküle und Anatomie im Konflikt? Die Systematik der Säugetiere. Stud. Integr. J. 20, 119-121.
|
|  |
