 |
Ursprung der Kolibris in der Alten Welt?
Neue Fossilfunde werfen Fragen zur Herkunft der Vogelfamilie auf
von André-A. Weller
Studium Integrale Journal
11. Jahrgang / Heft 2 - Oktober 2004
Seite 90 - 92
|
|
Einführung
Kolibris (Familie Trochilidae) werden in der mo-dernen Literatur als Schwestergruppe der Segler (Apodidae) und Baumsegler (Hemiprocnidae) klassifiziert und mit diesen in der Ordnung der Seglerartigen (Apodiformes) vereinigt (z.B. Sibley & Åhlquist 1990, Schuchmann 1999). Alle drei Gruppen teilen einige anatomische (u.a. Skelett des Flugapparates, Innervation der Flugmuskulatur, nur schwach ausgebildete Füße), physiologische (Malatdehydrogenase) und molekulare Gemeinsamkeiten (DNA-DNA-Rekombination), wobei Segler und Baumsegler im Körperbau und im Verhalten einander wesentlich stärker ähneln als den Kolibris. Insofern könnten die gemeinsamen Merkmale auch als Konvergenzen gedeutet werden, da Kolibris gegenüber beiden Seglergruppen weitere einzigartige Merkmalskomplexe besitzen. Aufgrund ihrer Fähigkeit zum Schwirrflug und einem extrem verlängerten Schnabel mit einer Kapillarzunge sind die kleinsten aller Vögel in der Lage, Nektarnahrung aus freistehenden Blüten aufzunehmen (Weller 1999). Von manchen Autoren (z.B. Wolters 1982) wurden sie deshalb früher in eine eigene Ordnung Trochiliformes gestellt.
|
|
Kolibrifossilien aus dem Oligozän?
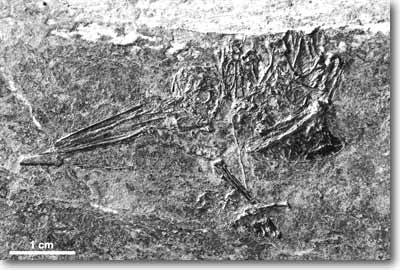 |
| Abb. 1: Eurotrochilus inexpectatus, ca. 2x so groß wie im Original. Foto: G. Mayr, Senckenberg-Museum Frankfurt a. M., Abdruck mit freundlicher Genehmigung. |
|
Mit etwa 330 Arten kommen Kolibris heutzutage ausschließlich in der Neuen Welt vor. Daß dies nicht immer so gewesen sein muß, deutet möglicherweise die Entdeckung und Klassifizierung der relativ gut erhaltenen fossilen Überreste zweier kolibriähnlicher Vögel aus einer Kalksteingrube im Baden-Württembergischen Frauenweiler an (Mayr 2004; Abb. 1). Sie werden auf ein Alter von 30-34 Millionen Jahren geschätzt und damit dem frühen Oligozän zugeordnet. Aufgrund des unerwarteten Auftretens in Europa und in Anlehnung an die der Vogelfamilie namensgebende Gattung Trochilus wurden die Vögel von ihrem Entdecker als „Eurotrochilus inexpectatus“ wissenschaftlich beschrieben. Die Funde aus den Archiven des Stuttgarter Naturkundemuseums gelten zumindest als ornithologische Überraschung, gibt es doch bisher nur fragmentarische Fossilbelege von Trochiliden aus dem karibischem Raum, die zudem wesentlich jünger datiert wurden (Quartär, ca. 1 Million Jahre; vgl. Schuchmann 1999).
Doch handelt es sich bei den süddeutschen Exemplaren tatsächlich um die nächsten Verwandten der heutigen Neuwelt-Kolibris? Immerhin weisen beide eine Reihe anatomischer Merkmale auf, die mit denen „moderner“ Arten Mittel- und Südamerikas durchaus vergleichbar sind. Auffällige Parallelen bilden vor allem die geringe Körpergröße (ca. 4-5 cm), der gegenüber dem Schädel mindestens um das 2,5fache verlängerte Schnabel (wenngleich die Spitze beim Holotypus, d.h. dem Referenzexemplar der neuen Gattung/Art, zu fehlen scheint und der Schnabel beim zweiten Exemplar gar nicht erhalten ist) sowie der Bau von Brustbein und Unterarm. Die Oberarm- und Unterarmknochen sind extrem verkürzt und kompakt gebaut, eine Einwölbung (Protrusion) am Oberarmkopf deutet eine erweiterte Drehbarkeit des Armgelenks an, die Eurotrochilus den Schwirrflug – ein einzigartiges Verhaltensmerkmal heutiger Kolibris – ermöglicht haben könnte. Übereinstimmungen bestehen auch in der Form des zum Schultergürtel gehörenden Rabenschnabelbeins (Coracoideum). Die Bein- und Fußknochen weisen sowohl Übereinstimmungen mit der Anatomie moderner Segler als auch der Kolibris (z.B. Hinterzehe) auf.
Dennoch unterscheiden einige Abweichungen im Körperbau Eurotrochilus von den modernen Kolibris. Hierzu zählen z.B. der Feinbau und die Längen der Mittelhand- und Fingerknochen sowie die Längenrelation von Brustbein und Elle. Außerdem scheint das Fossilexemplar einen zwar auffällig verlängerten, aber wesentlich kräftigeren Schnabel als Kolibris zu besitzen (Form in Relation zur Körpergröße etwa vergleichbar dem heimischen Eisvogel Alcedo atthis, mit mittlerer Schnabelhöhe von geschätzt > 3 mm; eventuell Erhaltungsartefakt?). Spekulativ bleibt zudem, ob Eurotrochilus überhaupt in der Lage war, vergleichbar den rezenten Trochiliden Nektar passiv durch eine Saugzunge aufzunehmen.
|
|
Fossile Bindeglieder zwischen Kolibris und Seglern?
Mayr (2004) ordnet die neue Gattung bzw. Art einer ursprünglichen Gruppe („stem group“) von Kolibris zu, die den heutigen Kolibris („crown group“) gegenübergestellt wird und zu der außerdem noch die von der Grube Messel bei Darmstadt bekannte Fossilart Parargornis messelensis (Mittleres Eozän) und die bislang nur im nördlichen Kaukasus gefundenen Argornis caucasicus (Oberes Eozän) und Jungornis tesselatus (Unteres Oligozän) gehören sollen (Mayr 2003b; vgl. Karhu 1999). Vertreter aller drei Gattungen zeigen z.T. ungewöhnliche anatomische Merkmalskombinationen und teilen einige Besonderheiten mit den Seglern und den modernen Kolibris. Jedoch ist teilweise unklar, inwieweit diese als gemeinsame abgeleitete Merkmale (abstammungsbedingt) oder als Konvergenzen zu werten sind (Mayr 2003b), da sich die betreffenden Arten in anderen Merkmalen signifikant von den rezenten Gruppen unterscheiden.
 |
| Abb. 2: Lesson-Amazilie (Amazilia amazilia); Lima, Peru. (Foto: André Weller) |
|
Beim einzigen als vollständiges Skelett vorlie-genden Fossilvogel dieser Gattungen, Parargornis, kontrastieren z.B. die Art der Befiederung, die Brei-te und Länge der Flügel mit Verkürzung der äuße-ren Handschwingen, die Länge der Beinknochen und die Ausbildung relativ kräftiger Füße relativ deutlich mit den korrespondierenden Merkmalen sowohl der Kolibris als auch der Segler (Mayr 2003b). Hingegen lassen die nur fragmentarisch erhaltenen Skelette von Argornis und Jungornis (nur Flügelelemente jeweils eines Individuums vorhanden) nur begrenzt Rückschlüsse auf deren Körperbau zu. Jungornis teilt mit Eurotrochilus und modernen Kolibris einige anatomische Elemente von wahrscheinlich funktioneller Bedeutung wie die stark, wenngleich nicht ganz so extrem verkürzten Oberarm- und Unterarmknochen und eine – allerdings abgeschwächte – distale Protrusion am Oberarm. Ein Unterscheidungsmerkmal von Jungornis ist das Fehlen einer verknöcherten Brücke am Kopf des Rabenschnabelbeins, die für rezente Vertreter der Kolibris charakteristisch ist. Weitere wichtige anatomische Merkmale wie z.B. die Schnabelform oder die Morphologie der Beine und Füße sind somit nicht vergleichbar. Eine aufgrund der Fundumstände nur die Merkmale der Flügel-knochen berücksichtigende phylogenetische Ana-lyse von Mayr (2003a) klassifiziert Jungornis als Schwestergruppe der Kolibris und Argornis als Schwestergruppe dieser beiden Taxa, woraus eine gemeinsame Abstammung aller drei Gruppen be-gründet wird. Das erst kurz danach beschriebene Taxon Parargornis wird in diesem Zusammenhang noch nicht mit aufgelistet, ist aber als nächstver-wandt mit Argornis einzustufen (Mayr 2003b). Im Gegensatz dazu ordnete Karhu (1999) Jungornis einer eigenen Familie der Jungornithidae zu, ohne jedoch die Trochilidae als Vergleichsgruppe zu berücksichtigen.
|
|
Vogel-Pflanze-Wechselbeziehungen
Einige Autoren nehmen an, daß Apodiden und Trochiliden monophyletisch sind, d.h. einen gemeinsamen Vorläufer besitzen, der als Insektenjäger durch einen kurzen Schnabel und kurze, breite Flügel (u.a. Parargornis) gekennzeichnet war (Cohn 1968; vgl. Mayr 2003a, b). Insofern wäre das Auftreten von kolibriartigen Vögeln wie Eurotrochilus, die möglicherweise bereits Nektar aufnehmen konnten, als durchaus revolutionär zu werten. Bisher wird angenommen, daß Vogel-Pflanze-Wechselbeziehungen (Nektarivorie, Ornithophilie) aufgrund ihrer relativ geringen Verbreitung im Vogelreich, noch dazu bei verwandtschaftlich nicht nahestehenden Gruppen – neben Kolibris sind hier u.a. die Nektarvögel, Honigfresser und Pinselzungenpapageien zu nennen – überwiegend in jüngeren erdgeschichtlichen Zeiträumen seit dem Tertiär mehrfach unabhängig voneinander entstanden sind (z.B. Grant 1994).
Die Tatsache, daß heute noch in der Paläotropis offenbar von fliegenden (als wichtige Unterscheidung zu sitzenden, nicht zum Schwirrflug fähigen) Blütenbesuchern bestäubte Pflanzen existieren, für die keine heute vorkommende Bestäuberart nachgewiesen werden kann (vgl. Westerkamp 1990), interpretiert Mayr (2004) als möglichen Hinweis darauf, daß diese Arten einst von Stammgruppen-Kolibris wie Eurotrochilus besucht wurden. Letztere könnten dann in der Folgezeit von den nicht vor dem Ende des Oligozän nachgewiesenen Singvögeln (Passeriformes) verdrängt worden sein.
Hinter diese Aussagen ist jedoch ein großes Fragezeichen zu setzen. Einerseits hätte gerade die postulierte Spezialisierung auf Nektarnahrung eine vorteilhafte ökologische Einnischung bedeutet und damit Konkurrenz mit anderen Vogelgruppen um Nahrungsressourcen vermieden, so wie es heute bei der Koexistenz von Kolibris, Singvögeln und anderen Vogelgruppen in weitaus größerer Artenfülle in Südamerika zu beobachten ist. Andererseits bleibt zweifelhaft, ob Koevolution mit ursprünglichen „Kolibris“ tatsächlich als Argument für das Auftreten von rezenten Pflanzenarten wie Canarina eminii (Campanulaceae) oder Impatiens sakeriana (Balsaminaceae) herangezogen werden kann (Mayr 2004), da deren Existenz über einen solch langen postulierten Zeitraum ohne ihren primären, längst ausgestorbenen Bestäuber zwangsläufig die Frage nach deren Überlebensfähigkeit aufwirft. Falls also tatsächlich kolibriähnliche Bestäuber im Oligozän in der Alten Welt existiert haben, wäre eine plausible Erklärung für deren Verschwinden wohl eher in grundlegenden klimatischen Veränderungen zu suchen. Andererseits könnten heute noch in Europa existierende ornithophile Pflanzen im Rahmen eines alternativen erdgeschichtlichen Deutungsmodells (vgl. Junker & Scherer 2001) einen wesentlich kürzeren als den postulierten Zeitabschnitt von ca. 30 Millionen Jahren seit dem Verschwinden der Vogelbestäuber andeuten.
|
Nektarivorie: Spezialisierung von Tierarten in der Ernährung auf Blütennektar. Ornithophilie: Ausbildung von Merkmalen bei Blütenpflanzen, die den Besuch von nektarfressenden Vögeln erleichtern. Konvergenz: mehrfach unabhängig (von verschiedenen Ausgangsstrukturen) entstandenes baugleiches Merkmal. rezent: heute lebend.
|
Ungeachtet der Herkunft der Vogel-Pflanze-Wechselbeziehungen, der Rolle ursprünglicher Bestäuber und deren Aussterben in Eurasien bleibt die Klassifikation von Eurotrochilus zur Familie Trochilidae umstritten, da Ähnlichkeiten im Körperbau nicht unbedingt als Indizien einer gemeinsamen Abstammung interpretiert werden können (Konvergenzproblem, vgl. Cracraft in Stokstad 2004). Aus biogeographischer Sicht erscheint es zudem fraglich, ob eine hypothetische Einwanderung durch eurasische Vorfahren heutiger Kolibris z.B. über die Beringstraße nach Nordamerika stattgefunden haben kann (Stokstad 2004), da sich das Entstehungs- und Ausbreitungszentrum moderner Trochiliden höchstwahrscheinlich in Südamerika befunden hat, wo etwa drei Viertel aller Arten anzutreffen sind. Nordamerika selbst ist von Kolibris höchstwahrscheinlich erst nach Schließung der mittelamerikanischen Landbrücke (datiert vor etwa 5 Millionen Jahren) besiedelt worden. Nicht zuletzt sind Merkmale und Verhaltensweisen, die nicht ohne weiteres aus der Fossilüberlieferung herangezogen werden können, wie Bioakustik und Brutbiologie, ebenso notwendige Indizien für eine korrekte Klassifizierung von bislang unbekannten Arten oder Gattungen; selbst wenn diese bekannt sind, ist die Zuordnung rezenter Vertreter zu einzelnen Vogelgruppen gelegentlich umstritten (Beispiel: Systematik bei Wasservögeln; vgl. Weller 2003).
|
|
Fazit
Der von Mayr (2004) benutzte Titel „Old World fossil record of moderntype hummingbirds“ scheint angesichts der vorliegenden Befunde zu Eurotrochilus verglichen mit heute lebenden Koli-bris ein wenig zu hoch gegriffen. Zudem erscheint angesichts der offensichtlich vorhandenen anato-mischen Merkmalsmosaike bei den hypothetischen Kolibri-Verwandten Jungornis, Argornis und Parargornis, die z.T. nur fragmentarisch vorliegen, und der vorhandenen Lücken in der Fossilüberlieferung eine Deutung von Gemeinsamkeiten dieser Fossilgruppen mit den heute vorkommenden For-men der Trochiliden als auch Eurotrochilus im Sin-ne von Abstammungsverwandtschaft (u.a. Mayr 2003b, 2004) als Spekulation. Als mögliches Binde-glied oder „missing link“ eignet sich Eurotrochilus kaum, da sich diese Form sowohl von den rezenten Kolibris als auch von postulierten Stammgruppenvertretern wie Jungornis in einigen Merkmalen unterscheidet. Der aktuelle Kenntnisstand rechtfertigt deshalb gleichermaßen die Annahme einer konvergenten Entstehung der genannten Gattungen bzw. Familien und damit eine Zuordnung zu verschiedenen Grundtypen (sensu Junker & Scherer 2001).
|
Literatur
- Cohn JMW (1968)
- The convergent flight mechanism of swifts (Apodi) and hummingbirds (Trochili). Ph.D. dis-sertation, University of Michigan, Ann Arbor.
- Grant V (1994)
- Historical development of ornithophily in the western North American flora. Proc. Natl. Acad. Sci. USA 91, 10407-10411.
- Junker R & Scherer S (2001)
- Evolution – ein kritisches Lehrbuch. 5. Auflage, Gießen.
- Karhu A (1999)
- A new genus and species of the family Jungornithidae (Apodiformes) from Late Eocene of the Northern Caucasus, with comments on the ancestry of hummingbirds. Smithson. Contrib. Paleobiol. 89, 207- 216.
- Mayr G (2003a)
- Phylogeny of early Tertiary swifts and hummingbirds (Aves: Apodiformes). Auk 120, 145-161.
- Mayr G (2003b)
- A new Eocene swift-like bird with a peculiar feathering. Ibis 145, 382-391.
- Mayr G (2004)
- Old World fossil record of modern-type hummingbirds. Science 304, 861-864.
- Schuchmann KL (1999)
- Family Trochilidae (Hummingbirds). In: Del Hoyo J, Elliot A & Sargatal J (eds) Handbook of Birds of the World. Vol. 5: Barn-owls to hummingbirds. Barcelona, pp 468-680.
- Sibley CG & Åhlquist JE (1990)
- Phylogeny and classificati-on of birds: a study in molecular evolution. New Haven, Connecticut.
- Stokstad E (2004)
- Bird evolution: Surprise hummingbird fossil sets experts abuzz. Science 304, 810-811.
- Weller AA (1999)
- Schillerfarben und Schwirrflug – Faszination Kolibris (1). Stud. Int. J. 6, 8-13.
- Weller AA (2003)
- Merkmalsmosaike bei Wasservögeln. Morphologische und molekulare Ähnlichkeiten passen nicht immer zusammen. Stud. Int. J. 10, 36-38.
- Westerkamp C (1990)
- Bird-flowers: hovering versus per-ching exploitation. Bot. Acta 103, 366-371.
- Wolters HE (1982)
- Die Vogelarten der Erde. Hamburg.
|
|  |
