 |
Wurde Europa doch früher besiedelt?
Überraschungen aus Ost und West von Sigrid Hartwig-Scherer
Studium Integrale Journal
9. Jahrgang / Heft 2 - Oktober 2002
Seite 67 - 73
|
 |
Zusammenfassung: Seit längerem gilt der europäische Homo sapiens als Abkömmling eines „Afrikaners“, der vergleichsweise spät – zwischen 100.000-200.000 radiometrischen Jahren – den schwarzen Kontinent verlassen hat. Bei seiner Einwanderung soll er alle ansässigen europäischen Formen verdrängt haben. Die früheren europäischen Formen wie z.B. Homo heidelbergensis hatten dieser Vorstellung gemäß Europa auch nicht vor 0,5 Millionen radiometrischen Jahren (MrJ) besiedelt, denn man nahm bis vor einigen Jahren an, daß Homo ergaster/erectus erst nach der Entwicklung der Kulturstufe des Acheuléen und dem damit verbundenen angeblich höheren kulturellen Niveau sein Ursprungsland Afrika hätte verlassen können. Deshalb galten alle Fundstätten Europas, die die Altersmarke von 0,5 MrJ überschritten, als zweifelhaft. Neuere, zwischen 1,8 und 0,8 MrJ alte Fundstätten an zwei entgegengesetzten Enden Europas – Georgien und Spanien – ziehen nun die Vorstellung eines anthropologisch jungen Europas in Zweifel und offerieren als mögliche Alternative, daß auch der europäische Homo sapiens wesentlich ältere Wurzeln innerhalb Europas haben könnte.
Neue DNA-Studien lassen ebenfalls die späte und „reine“ Out-of-Africa-Hypothese fragwürdig erscheinen. Danach scheint der Mensch in mehreren Wellen nach Europa eingewandert zu sein, sich mit den jeweils ansässigen Formen vermischt und vor Ort weiter verändert zu haben. Diese Vorstellung entspricht auch dem Grundtypmodell: die aus dem afrikanisch-levantinischen Korridor auswandernde polyvalente Basisform Homo veränderte sich aufgrund von Mikroevolutionsprozessen (Anpassung), Wanderungen und Hybridisierung und bildete recht unterschiedliche Formen und/oder Arten aus, deren Entstehung mittels eines linearen Entwicklungsmodells bislang von der Paläanthropologie nicht befriedigend erklärt werden konnte.
| |
|
Wie afrikanisch ist der europäische Homo sapiens wirklich? Eine kurze Einführung
Bislang war Europa für die Paläanthropologie bestenfalls ein Cul-de-Sac (Sackgasse), ein abgestandener Altwasserarm, in dem sich wenig erfolgreiche menschliche Formen „stauten“ und nur solange überdauerten, bis der überlegene afrikanische Homo sapiens sie – allerdings erst relativ spät – vollständig eliminierte. So zumindest liest sich die gängige Version seit den 1980er Jahren. Dieses Späte Monozentrische Modell (1. Kasten), auch „Out-of-Africa-II-Modell“ genannt, ist weiterhin (noch) die populärste Hypothese zur Entstehung des anatomisch modernen Menschen: der vergleichsweise spät in Afrika entstandene anatomisch moderne Mensch bildete nach diesem Modell praktisch ausschließlich die genetische Grundlage des europäischen Homo sapiens. Auf dieser Grundlage wurden alle europäischen Fundkomplexe, die älter als 0,5 MrJ datiert wurden, sehr kritisch betrachtet und meist als Fehldatierungen verworfen. Doch seit einigen Jahren gewinnt das alte Europa (d.h vor 0,5 MrJ) zunehmend paläanthropologische Bedeutung: so warten an den beiden entgegengesetzten Enden Europas zwei herausfordernde Fundkomplexe darauf, in das bestehende Modell integriert zu werden oder es umzustoßen: am östlichen Tor Europas, in Georgien, begehrt die sehr frühe Menschenform Homo ergaster/erectus Einlaß und in Spanien erhebt eine ganze Palette unterschiedlich alter Menschenfunde den Anspruch auf die Vorfahrenschaft verschiedener europäischer Menschen-Formen. Im Zuge dieser Entwicklung wurden auch viele der früher als „zu alt“ geltenden Fundorte rehabilitiert. So diskutiert man die Möglichkeit, ob die Funde in Orce, Südspanien, doch zwischen 1,2 und 1,8 MrJ alt sein könnten und zusammen mit dem georgischen Dmanisi (1,75 MrJ) zu den ältesten menschlichen Spuren Europas gehören. Auch die anderen alten Fundstellen gelten damit als Zeugnis für eine frühe Besiedlung Europas (CULOTTA 2001; BALTER 2001; GIBBONS 2001).
Drei gängige Hypothesen zur Entstehung
des Homo sapiens und das Grundtypmodell
|
|
| 1. Das Späte Monozentrische Verdrängungs-Modell (Out-of-Africa II)
Neben der Fossilverteilung sind DNA-Befunde über stark verarmte Variabilität die Hauptargumente für diese Vorstellung: der anatomisch-moderne Mensch habe sich erst vor relativ kurzer Zeit (vor maximal 200.000 rJ) und nur an einem Ort entwickelt. Hier wird in den meisten Fällen Afrika angeführt, manchmal auch der Nahe Osten (siehe z.B.WADDLE 1994). Von dort aus habe er den Rest der Welt besiedelt und alle älteren ansässigen Formen vollständig verdrängt. Europa war damit vollständig vom Schauplatz der menschlichen Evolution verschwunden und konservierte bestenfalls ausrangierte „Modelle“, die sich in Europa etwas länger halten konnten, schließlich aber doch erfolgreicheren afrikanischen Gruppen weichen mußten.
2. Das Multiregionale Modell (Out of everywhere)
Das ältere, aber momentan (noch?) weniger populäre Multiregionale Modell (THORNE & WOLPOFF 1992) geht von einer lokalen Evolution aus. Danach sei der heutige Mensch mehrfach und im wesentlichen unabhängig voneinander in Afrika, Asien, Europa aus regional ansässigen H. erectus- oder ergaster-ähnlichen Vorfahren entstanden. Dabei werden die DNA-Befunde, die von den meisten Genetikern eher mit einem rezenten Ursprung und einem Ausbreitungszentum (Evolution nach einem „Flaschenhalsereignis“) erklärt werden, anders interpretiert. Auch der zweite Einwand, daß die Entstehung an drei verschiedenen Orten, d.h. weitgehend unabhängig relativ unwahrscheinlich sei, wird aufgrund ihrer nicht-cladistischen, sondern
|
gradualistischen Artvorstellung als unproblematisch abgewiesen (für einen Multiregionalisten sind H. sapiens und erectus nur Chronospecies, d.h. im wesentlichen nur eine sich über die Zeit verändernde Art).
3. Die afro-europäische sapiens-Hypothese (Multiregionales Out-of-Africa-Modell)
Vor Jahren wurde dieses Modell von einem Deutschen vorgeschlagen (BRÄUER 1984). Als „Kompromiß“ der beiden obigen Modelle wurde es in der Süddeutschen Zeitung (Nr. 72, 26. 3. 2002) als Multiregionales Out-of-Africa-Modell bezeichnet. Den mittelpleistozänen europäischen (und entsprechend den asiatischen) Formen wird ein gewisser genetischer Beitrag an der Evolution des modernen Menschen zugestanden, d.h. Hybridisation der ansässigen Formen mit den vielleicht sogar mehrfach aus Afrika zuwandernden Menschen wird für möglich gehalten.
4. Grundtypmodell
Dieses Modell vereint verschiedene Aspekte der obigen drei gängigen Hypothesen: es geht von umfänglichen Wanderungen, in-situ-Mikroevolution und einer genetischen Verträglichkeit = Hybridisierung aller Mitglieder eines Grundtyps aus (Vermischung von ansässigen und zuwandernden Formen). Die Vorstellung zu den absoluten Zeiten ist eine grundlegend andere und das Entstehungszentrum wird eher im Levantinischen Korridor vermutet. Als Ausgangsformen der jeweiligen Grundtypen werden poylvalente Stammformen postuliert.
|
|
|
|
Ältester Türsteher Europas – kam der erste „Europäer“ aus dem Kaukasus?
Als man vor über 10 Jahren am östlichsten Ende Europas in Dmanisi (Georgien; Abb. 1), die ersten Spuren menschlicher Besiedlung entdeckte, war das schon eine Sensation für sich (GABUNIA & VEKUA 1995). Dieser Erstfund, ein Unterkiefer, hatte Ähnlichkeiten mit asiatischen Homo erectus-Formen, aber auch mit dem afrikanischen Homo ergaster, die einige Forscher zwischenzeitlich auch wieder zu einer Art vereinigen wollen (ASFAW et al. 2002). Als dann klar wurde, daß es sich bei diesem Unterkiefer um eine weit über 1 MrJ alte Form handelt, galt der Fund bald als eines der ältesten Vorkommen der Gattung Homo außerhalb Afrikas und potentieller Eroberer Europas und Asiens (DEAN & DELSON 1995). Im Laufe der mehrere Jahre andauernden Grabungen kamen an der Fundstelle südlich von Tbilisi noch zwei partielle Schädel von einem jungen Mädchen und einem Jugendlichen hinzu (GABUNIA et al. 2000), deren Gesichtsmorphologien stark dem afrikanischen Homo ergaster aus Koobi Fora, Kenya, ähneln. Beide Fundkomplexe wurden mit einem ähnlichen geologischen Alter eingestuft (ca. 1,8 für Koobi Fora und ca 1,75 MrJ für Dmanisi). Die ersten Besiedler Eurasiens besaßen also genetische Verbindungen zu den afrikanischen und den asiatischen Menschenformen (BALTER & GIBBONS 2000). GABUNIA und Mitarbeiter nehmen an, daß sie den Kaukasus vom Levantinischen Korridor her erreichten, über den es einen regen Faunenaustausch zwischen Asien und Afrika gab (GABUNIA et al. 2000; Abb. 1).
Der Lebensraum in Dmanisi muß fast paradiesisch gewesen sein (möglicherweise auch für andere Primaten, wie neueste Funde zeigen; vgl. HARTWIG-SCHERER 2002): Das Mosaik von offener Steppe und Galeriewäldern mit vielfältigen Ressourcen und einem angenehm warmen Klima bot zumindest eine gute Ausgangslage für weitere Eroberungen in angrenzende Gebiete in Ost-Europa und West-Asien. Der Dmanisi-Mann fertigte jedoch nicht wie seine späteren afrikanischen Artgenossen Acheuléen-Faustkeile an, sondern „nur“ Werkzeuge aus der core-chopper Steinwerkindustrie, also im wesentlichen einfache Abschläge. Damit kann nach den Ausgräbern ein Fortschritt in der Steinwerkzeug-Technologie nicht die treibende Kraft für die Wanderungen gewesen sein, sondern möglicherweise eher die Expansion des Lebensraums aufgrund des zu vergrößernden Jagdreviers: Die Menschen zogen gleichermaßen ihren Beutetieren hinterher. Die noch Ende des letzten Jahrhunderts gültige Annahme, daß der Mensch erst nach der Entwicklung des Acheuléen-Fundkomplexes, die Fähigkeit besaß, Afrika zu verlassen (d.h. nach 1 MrJ ) und angrenzende Kontinente zu erorbern, wurde zuvor schon durch die Neudatierung des Modjokerto-Kindes, Java, auf 1,8 MrJ (SWISHER et al. 1994) in Frage gestellt.
Die Beschreiber (GABUNIA et al. 2000) sehen in ihren alten Funden nicht nur Merkmale früher afrikanischer und asiatischer Formen, sondern auch fortschrittliche Züge, die auf spätere Evolutionsstufen des Menschen hinweisen sollen. Da ist der Schritt nicht mehr allzu weit, den Dmanisi-Menschen zum Vorläufer späterer europäischer Mittelpleistozänformen und schließlich zum anatomisch modernen Menschen zu machen (BRÄUER & SCHULTZ 1996; ROSAS & BERMUDEZ DE CASTRo 1998). Diese Vorstellung wäre jedoch dem vorherrschenden Späten Out-of-Africa-Modell gegenläufig.
|
|
... oder aus Spanien? Neues vom Westen
Gegen dieses Modell scheinen auch spanische Funde aus Atapuerca zu sprechen. Dort fand man eine ganze Fundserie unterschiedlich alter menschlicher Fossilien und Besiedlungsspuren über einen enorm langen Zeitraum von fast 0,75 MrJ hinweg. Die Fossilien zeichnen sich durch eine hochvariable Morphologie aus, die fast alle bekannten menschlichen Merkmale in sich vereint. Damit bietet sie sich – nach Meinung der Beschreiber – als ideale Ausgangspopulation für alle späteren Formen an. Ein Forscher kommentiert in Science: „Nach Atapuerca müssen wir alles neu überdenken!“ (BALTER 2001)
Die ältesten Fossilien, die im Höhlensystem Gran Dolina bei Atapuerca gefunden wurden, datieren die Forscher auf mehr als 0,78 MrJ. Sie kommen – ähnlich wie die um 1 MrJ älteren Dmanisi-Menschen und deren asiatischen Zeitgenossen – nur mit einfachen Abschlagwerkzeugen vor (CARBONELL et al. 1995). Eric DELSON wundert sich, wieso es keine fortschrittlicheren Werkzeuge gab: „Haben sie einfach nur alte Bräuche auf dem Weg verloren?“ Die Atapuerca-Formen gehören ohne Zweifel mit zu den ältesten Bewohnern Europas (GUTIN 1995). Aufgrund der morphologischen Besonderheit wurde mit Hilfe des älteren Materials (um 0,7 MrJ) die neue Art Homo antecessor errichtet (BERMUDEZ DE CASTRo et al. 1997), die jedoch nicht von allen Anthropologen als gültige Art anerkannt wird. Nach Aussagen der Finder soll die neue Art der Vorfahr aller europäischen Formen sein, d.h. für H. heidelbergensis, für den anatomisch modernen Menschen und für den Neandertaler (siehe auch 2. Kasten). Eine solche Vorstellung ist völlig unvereinbar mit der gängigen späten monozentrischen Entstehung in Afrika (1. Kasten).
Die jüngeren Formen, die aus dem Höhlensystem Sima des los Huesos bei Atapuerca um 300.000 rJ stammen, werden von ihren Beschreibern als Homo heidelbergensis klassifiziert. Diese Formen haben nach ARSUAGA et al. (1993) mehr oder weniger große morphologische Ähnlichkeiten mit Formen aus Steinheim, Petralona, Arago, Bilzingsleben, Neandertal (Abb. 1 und 2), aber auch mit Afrikanischen Formen wie Bodo und Broken Hill (letztere werden von manchen Forschern auch als H. rhodesiensis bezeichnet und als afrikanisches Pendant des europäischen H. heidelbergensis angesehen). Die spanischen H. heidelbergensis-Formen werden als hochpolymorph (vielgestaltig) beschrieben, wobei die Intensität der Neandertalmerkmale mit der Zeit stetig zunähme (d.h. die geologisch jüngsten Populationen zeigen die ausgeprägtesten Neandertalzüge) (ARSUAGA et al. 1993; CARBONELL & RODRIGUEZ 1994). Somit hat der Fossilkomplex aus Atapuerca auch Konsequenzen für die Neandertalevolution (RIGHTMIRE 1997; vgl. 2. Kasten).
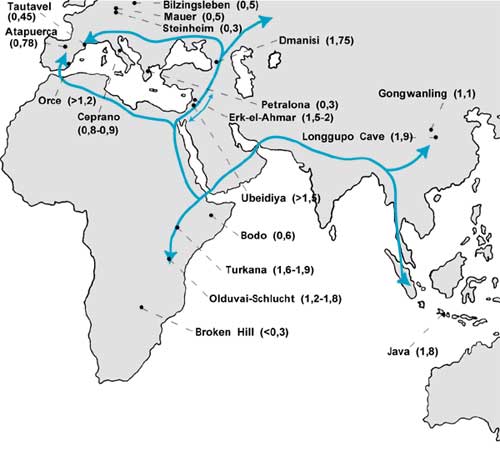 |
| Abb 1: Die geographische und geologisch-zeitliche Verteilung der wichtigsten Fundorte fossiler Formen der Gattung Homo. Die Pfeile geben mögliche Ausbreitungsrichtungen an. Die zeitliche und geographische Verteilung dieser Homo-Arten ist unterschiedlich gut verträglich mit den drei grundsätzlich verschiedenen Modellen zur Evolution des Homo sapiens (siehe 1. Kasten). |
|
Den ehrgeizigen Vorstellungen der spanischen Ausgräber folgend hätte hier also eine mehr oder weniger kontinuierliche in situ-Evolution (eine sich vor Ort vollziehende evolutive Veränderung) stattgefunden, und zwar von Homo antecessor über H. heidelbergensis zu H. neanderthalensis einerseits und zu H. sapiens andererseits.
Die Beschreiber schlußfolgern aus den Merkmalen von Homo antecessor, daß sowohl H. sapiens als auch H. neanderthalensis gemeinsame und sehr alte Wurzeln haben sollen – und zwar in Europa. Sie glauben nicht, daß sich die beiden Linien – H. sapiens und H. neanderthalensis – schon früh in Afrika trennten und erst spät und nacheinander nach Europa einwanderten, wie es einige DNA-Studien (siehe HARTWIG-SCHERER 2000) nahelegen. Sie glauben, daß H. antecessor via Golf von Gibraltar über Spanien nach Europa eingewandert ist.
Der südspanische Fund bei Orce (zwischen 1,2 und ?1,8 MrJ geschätzt; GIBERT et al. 1998; MARTINEZ NAVARRO et al. 1997; MOYA-SOLA & KOHLER 1997) und die ältesten Atapuerca-Spuren (ROSAS & BERMÚDEZ DE CASTRO 1999), die ebenfalls an die 1 MrJ-Marke herankommen, werden zusammen mit den anderen europäischen Funden als ein Indiz für die Besiedlung Europas von Südwesten aus gewertet und nicht, wie die georgischen Kollegen vorschlagen, aus dem Osten. Die zeitlich-geographische Abfolge dieser Formen innerhalb Europas – Ceprano (Italien, ca. 0,8 MrJ), Isernia (Italien, 0,7 MrJ), Mauer (Deutschland, >0,5 MrJ), Bilzingsleben (Deutschland, >0,5 MrJ), Boxgrove (England, 0,5 MrJ), Schoeningen (Deutschland, 0,4 MrJ), Arago (Frankreich, 0,4 MrJ) und Petralona (Griechenland, 0,3 MrJ) (Abb. 1) – sollen die Besiedlungsroute Nordafrika-Gibraltar-Spanien-Europa belegen, da die ältesten Funde nicht im Osten, sondern im tiefsten Westen zu finden sind. Nach SCHRENK (2000) ist eine frühe Besiedlung über Gibraltar gut vorstellbar, begründet vor allem wegen faunistischer Ähnlichkeiten zwischen Marokko und Südspanien. Wegen des schwankenden Meeresspiegels war dieser Sund nicht immer – wie heute – 13, sondern manchmal nur 6 km breit, was ein Übersetzen ermöglicht haben könnte. Die alten spanischen Fundstätten werden deswegen auch mit denen Nordwestafrikas in Verbindung gebracht und als Wurzeln der europäischen Variante des Homo erectus (den die spanischen Kollegen Homo antecessor nennen) diskutiert.
| Klicken Sie auf einen Schädel um ihn gross zu sehen |
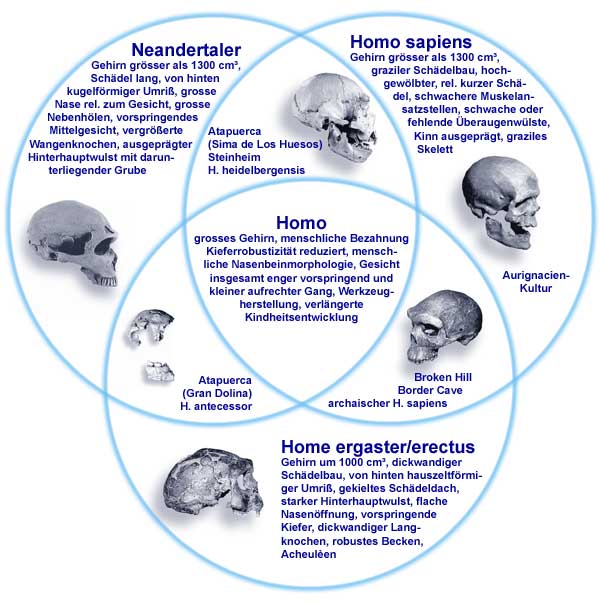 |
| Abb. 2: Die drei morphologischen Hauptgruppen des Menschen und die mittelpleistozänen Mischformen mit den jeweils charakteristischen Merkmalen. Funde und Arten, die spezielle Merkmale von zwei (oder sogar drei) Gruppen in sich vereinigen, sind in den entsprechenden Überlappungszonen anzusiedeln. Homo antecessor weist Merkmale aller drei Hauptgruppen auf; in der Abbildung ist dies nicht genau darstellbar. (Nach JUNKER & SCHERER 2001; nach STRINGER) |
|
Unterstützt durch verschiedene DNA-Studien stellen dagegen die Anhänger der Späten Monozentrischen Hypothese die Modernität der „sapiens-ähnlichen“ Merkmale bei Homo antecessor (BERMÚDEZ DE CASTRO et al. 1999) in Frage, da diese Form in ihren Augen dafür um fast mehr als eine halbe MrJ zu alt sind. Allerdings sprechen andere DNA-Studien gegen den späten und ausschließlich afrikanischen Ursprung und eher für eine mehrfache und relativ frühe Besiedlung (TEMPLETON 2002).
Zweifel an der Interpretation von bestimmten Morphologien gibt es auch für die späteren Funde: Das Axialskelett des Homo heidelbergensis aus Atapuerca gleicht dem modernen Menschen mehr als denen der anderen mittel- und oberpleistozänen Formen (CARRETERO et al. 1999). Entweder man hält nun diese Grazilität für ein modernes Merkmal, das sehr früh auftritt. Dann muß entweder das Alter und/oder der Evolutionsmodus des H. sapiens stark verändert werden, oder man muß das Alter der Atapuercafossilien anzweifeln (was in diesem Fall eher schwierig ist). Oder aber man hält diese Modernität gar nicht für eine echte Modernität, d.h. Neuerwerbung des sapiens, sondern betrachtet es als plesiomorphes Merkmal (d.h. von früheren Formen übernommen). Dann aber fällt ein mächtiger Grundpfeiler zur Artdefinition von H. sapiens (Abb. 2) in sich zusammen, was weitere schwerwiegende Konsequenzen hätte. Die Anthropologie steht vor der schwierigen Entscheidung, ob man diesen diagnostisch signifikanten Verlust in Kauf nehmen will, nur um die Späte Out-of-Africa-Hypothese zu retten. Ein Vorschlag TEMPLETONs, seine DNA-Studien als Indiz für eine einzige Art oder Rasse der Gattung Homo zu interpretieren (TEMPLETON 1998), stieß auf heftige Kritik, auch wenn dadurch obige Probleme scheinbar weniger schwer wiegen. Friedemann SCHRENK schlägt als Kompromiß eine „multiregionale Out-of-Africa-Hypothese“ vor – für manche Paläanthropologen weder Fisch noch Fleisch. Eine Einigkeit besteht unter den Anthropologen jedenfalls nicht.
Gehört der Neandertaler nun doch zu unseren Vorfahren?
|
|
| In den vergangen Jahren hat sich das Bild vom Neandertaler und seiner Verbindung mit uns mehrfach grundlegend geändert. Die beträchtlichen Unterschiede in der mitochondrialen DNA (mtDNA) des Neandertalers im Vergleich zu Homo sapiens wurden dahingehend interpretiert, daß es sich um zwei sehr früh getrennte Linien handelt (ca. 600-800.000 rJ), die auch gemäß dem Späten Monozentrischen Modell keine genealogische und genetische Verbindungen miteinander gehabt haben können. Das angeblich sehr hohe Alter der Neandertallinie, das aus DNA-Vergleichen von Neandertalern und heutigen Menschen gefolgert wurde (KRINGS et al. 1999; KRINGS et al. 1997; OVCHINNIKOV et al. 2000; HARTWIG-SCHERER 2000), schoben ihn erst einmal auf einen fernen Seitenzweig.
Eine australische Arbeitsgruppe (ADCOCK et al. 2001) kritisierte diese Schlussfolgerung berechtigterweise mit dem Einwand, daß keine gleichalten H. sapiens-Formen als Kontrollgruppen verwendet wurden. Sie unternahmen den DNA-Vergleich mit einem frühen sapiens-Fund von Lake Mungo (LM3, Australien), dessen Alter auf umstrittene 62.000 rJ geschätzt wird (was an sich schon eine Sensation ist: es ist das älteste Begräbnis, bei dem das Bestreuen mit rotem Ocker nachgewiesen wurde). Die DNA-Untersuchung ergab, daß bei dieser australischen sapiens -Form ebenfalls eine mtDNA-Linie vorhanden ist, die bei heutigen Menschen – also bei der gleichen Art – nicht mehr vorkommt! Wenn also eine Linie innerhalb des anatomisch modernen Menschen, zu der LM3 gehört, ausgestorben ist, dann kann auch etwas ähnliches mit der mtDNA von Neandertalern passieren, wie ein Kommentar feststellt (RELETHFORD 2001). Ähnlich klingt die Schlußfolgerung in der neuesten mtDNA-Studie (HAWKS & WOLPOFF 2001) von dem auf 28.000 rJ datierten Neandertaler von Vindiya (SMITH et al. 1999), der ähnlich dem Kind von Lagar Velho 1 (Portugal) auch ein Hybrid zwischen den beiden Menschenformen gewesen sein könnte (vgl. HARTWIG-SCHERER 2000). HAWKS & WOLPOFF (2001) folgern, daß die Fossilform aus der Mezmaiskaya-Höhle (OVCHINNIKOV et al. 2000), deren mt-DNA eine alte Linie nahelege, in Wirklichkeit gar kein Neandertaler gewesen sein soll. Man könne diese Gruppen genauso gut als Mischpopulationen einer einzigen großen Spezies mit sapiens und neanderthalensis-Merkmalen ansehen, deren mtDNA aufgrund von natürlicher Selektion substituiert worden ist – ganz im Sinne der Multiregionalhypothese und kompatibel mit dem Grundtypmodell.
Neuere Daten geben zudem Hinweise auf eine außerordentlich hohe Evolutionsgeschwindigkeit der mtDNA (Übersicht in LOEWE & SCHERER 1997). Danach müßte eigentlich die mtDNA-Uhr völlig neu geeicht werden und kann nicht als Grundlage für solche weitreichenden Rückrechnungen verwendet werden.
Die Frage wird wieder neu gestellt, ob zwischen uns und den Neandertalern doch eine gewisse genetische Kontinuität gegeben ist. Sollte der abgeschobene Neandertaler doch unter unseren Vorfahren zu finden sein? Zumindest müssen die drei theoretisch möglichen Positionen des Neandertalers und die jeweils zugrundeliegende Hypothesen (siehe 1. Kasten) neu bedacht werden: a) direkter Vorfahre (Multiregionale Hypothese), b) Seitenzweig ohne jede genetische Beteiligung (Späte Monozentrische Hypothese), oder c) genetische Mitbeteiligung (Afroeuropäische sapiens-Hypothese = Multiregionale Out-of-Africa-Hypothese).
Inzwischen müssen wir uns verwundert mit der Frage auseinandersetzen: Ist der Neandertaler deswegen nicht unser Vorfahre, weil er einer zu alten Linie angehörte oder gehört er gerade wegen seines Alters mit zu unseren Vorfahren?
|
|
|
|
... oder beides? Ein möglicher Konsens
Auf einen Konsens konnte sich die Paläanthropologie einlassen, nämlich daß ab 1,9 MrJ an allen Enden der Alten Welt (Afrika, Eurasien, Südostasien und Ost-Europa) erste menschliche Besiedlungsspuren auftauchten. Ursprünglich ging man davon aus, daß H. erectus/ergaster erst lange Zeit nach seiner Entstehung aus Afrika auswanderte und die Alte Welt besiedelte. Doch da Afrika in den letzten Jahren sukzessiv seinen langen zeitlichen Vorsprung eingebüßt hat, kommen auch angrenzende Kontinente für die Wiege der Menschheit in Betracht. So wird sogar schon bei dieser ersten Evolutionsetappe der mögliche Entstehungsort auf den afroasiatischen Raum ausgeweitet: Gerhard BOSINSKI vom Museum Monrepos in Neuwied, einer der Mitbeschreiber von Dmanisi, vermutet, daß sich die Entwicklung des Menschen in einer Region vollzogen habe, die sich von Ostafrika bis ins südliche Eurasien erstreckte (Abb. 1). Der Levantinische Korridor, das heutige Israel mit angrenzenden Staaten, verbindet Europa, Asien und Afrika und ist aufgrund eines etwa 1,9 MrJ alten Fundorts in Nordisrael (Erk-el-Ahmar; HAGAI & LEVI 2001) nicht nur ein möglicher Durchgangs-, sondern vielleicht auch Entstehungsort dieses ersten echten Menschen (H. erectus/ergaster).
 |
| Die unterschiedlichen Merkmalsmosaike lassen sich kaum in eine evolutiv-lineare Abfolge bringen |
 |
|
Was die Entstehung des Homo sapiens, also des anatomisch modernen Menschen betrifft, ist bislang überhaupt keine Übereinstimmung in Sicht. Seit den 1980er Jahren galt mehrheitlich die Vorstellung, daß der anatomisch moderne Mensch erst relativ spät (um 0,2 MrJ) in Afrika entstanden ist (1. Kasten), die Alte Welt besiedelt und alle anderen ansässigen Formen vollständig verdrängt hat. Das „europäische“ Alternativmodell postuliert nun, daß ein afrikanischer Vorfahre wesentlich früher (vor 1 MrJ) nach Europa eingewandert ist. Ob die Besiedlung über die Route Nordafrika – Gibraltar oder Levantinischer Korridor-Kaukasus oder beides erfolgte, bleibt offen (Abb. 1).
 |
| Abb. 3: Schädel eines australischen Ureinwohners. (Staatliches Museum für Naturkunde, Karlsruhe) |
|
Die verwirrende Vielgestaltigkeit der mittelpleistozänen Formen machen den Paläanthropologen seit jeher Kopfzerbrechen, was man auch in den uneinheitlichen taxonomischen Bezeichnungen erkennen kann. Ein und dieselbe europäische mittelpleistozäne Form kann man in der Literatur schon mal als europäischer Homo erectus, als H. heidelbergensis oder als früher archaischer H. sapiens klassifiziert finden. Sie sind ein einzigartiger Schmelztiegel unterschiedlicher Morphologiemosaike aus erectus-, heidelbergensis-, neanderthalensis- und sapiens-anmutenden Merkmalen, die sich kaum in eine evolutiv-lineare Abfolge bringen lassen. Das „make-love-not-war“-Modell mit mehreren Besiedlungswellen könnte schon eher dieses bunte Netzwerk an Morphologien beschreiben.
Das Grundtyp-Modell erklärt die nichtlineare morphologische Heterogenität innerhalb aller Mitglieder eines Grundtyps generell mit der Polyvalenz (genetische Vielseitigkeit) der Ausgangsform. Danach ist es nicht ungewöhnlich, wenn einzelne Merkmale späterer Formen – hier beim Grundtyp Homo z.B. die Art H. sapiens – auch schon bei frühen Formen – z. B. bei Homo antecessor oder H. heidelbergensis – nachweisbar sind, ohne daß man eine (genealogische) Direktverbindung zwischen diesen zugrunde legen muß: die für spätere Arten typischen Merkmale können sporadisch zeitlich und räumlich weit entfernt auftreten und wieder verschwinden. Mikroevolution, genetische Drift und Selektion können vorher vorhandene Merkmale (die latent vorhanden waren oder nur sporadisch auftauchen) als (Art)definierende Kombinationen fixieren (Artbildung).
Da man im Grundtypmodell von ausgiebigen Wanderungen ausgeht, ist eine mehrfache Besiedlung Europas – sowohl über Gibraltar als auch über den Levantinischen Korridor – denkbar und sogar wahrscheinlich. Nach diesem Modell könnten sich die pleistozänen Homo-Formen lokal zu verschiedenen Arten ausdifferenziert und in unterschiedlicher Intensität mit später einwandernden sapiens-ähnlichen Formen vermischt haben. Eine solche Bevölkerungsmischung ist in diesem Modell zu erwarten, zumal Hybridisierung das wichtigste Kriterium für die Zugehörigkeit zu einem bestimmten Grundtyp ist. Sowohl die spanischen als auch die georgischen Menschenformen sind nach dem Grundtypmodell verschiedenartige Ausdrucksformen des gleichen Grundtyps, und partizipieren auf diese Weise am Genpool heutiger Menschen, ohne direkte Vorfahren sein zu müssen. Deshalb sind auch die „sapiens“-Züge mancher früher Formen nicht allzu verwunderlich, ebensowenig wie manch ursprünglich anmutende Merkmale heutiger Menschenpopulationen (z. B. australische Ureinwohner; Abb. 3).
|
Literatur
- ADCOCK GJ, DENNIS ES, EASTEAL S, HUTTLEY GA, JERMIIN LS, PEACOCK WJ & THORNE A (2001)
- Mitochondrial DNA sequences in ancient Australians: Implications for modern human origins. Proc. Natl. Acad. Sci. USA 98, 537-542.
- ARSUAGA J-L, MARTINEZ I, GARCIA A, CARRETERO J-M & CARBONELL E (1993)
- Three new human skulls from the Sima de los Huesos Middle Pleistocene site in Sierra de Atapuerca, Spain. Nature 362, 534-537.
- ASFAW B, GILBERT WH, BEYENE Y, HART WK, RENNE PR, WOLDEGABRIEL G, VRBA ES & WHITE TD (2002)
- Remains of Homo erectus from Bouri, Middle Awash, Ethiopia. Nature 416, 317-320.
- BALTER M (2001)
- News: In search of the first Europeans. Science 291, 1722-1725.
- BALTER M & GIBBONS A (2000)
- Paleoanthropology. A glimpse of humans’ first journey out of Africa. Science 288, 948-950.
- BERMUDEZ DE CASTRO JM, ARSUAGA JL, CARBONELL E, ROSAS A, MARTINEZ I & MOSQUERA M (1997)
- A hominid from the lower Pleistocene of Atapuerca, Spain: possible ancestor to Neandertals and modern humans. Science 276, 1392-1395.
- BERMÚDEZ DE CASTRO JM, ROSAS A, CARBONELL E, NICOLÁS ME, RODRÍGUEZ J & ARSUAGA JL (1999)
- A modern human pattern of dental development in Lower Pleistocene hominids from Atapuerca-TD6 (Spain). Proc. Natl. Acad. Sci. USA 96, 4210-4213.
- BRÄUER G (1984)
- Präsapiens-Hypothese oder Afro-europäische Sapiens-Hypothese? Z. Morphol. Anthropol. 75, 1-25.
- BRÄUER G & SCHULTZ M (1996)
- The morphological affinities of the Plio-Pleistocene mandible from Dmanisi, Georgia. J. Hum. Evol. 30, 445-481.
- CARBONELL E, BERMUDEZ DE CASTRO JM, ARSUAGA JL, DIEZ JC, ROSAS A, CUNCA-BESCOS G, SALA R, MOSQUERA M & RODRIGUEZ XR (1995)
- Lower Pleistocene hominids and artifacts from Atapuerca-TD6 (Spain). Science 269, 826-830.
- CARBONELL E & RODRIGUEZ XR (1994)
- Early middle Pleistocene deposits and artefacts in the Gran Dolina site (TD4) of the Sierra de Atapuerca (Burgos, Spain). J. Hum. Evol. 26, 291-311.
- CARRETERO JM, LORENZO C & ARSUAGA JL (1999)
- Axial and appendicular skeleton of Homo antecessor. J Hum Evol 37, 459-499.
- CULOTTA E, SUDGEN A & HANSON B (2001)
- Humans on the Move. Science 291, 1721.
- DEAN D & DELSON E (1995)
- Homo at the gates of Europe. Nature 373, 472-473.
- DISOTELL TR (1999)
- Human evolution: origins of modern humans still look recent [see comments]. Curr. Biol. 9, R647-R650.
- GABUNIA L & VEKUA A (1995)
- A Plio-Pleistocene hominid from Dmanisi, East Georgia, Caucasus. Nature 373, 509-512.
- GABUNIA L, VEKUA A & LORDKIPANIDZE D (2000)
- The environmental contexts of early human occupation of Georgia (Transcaucasia). J. Hum. Evol. 38, 785-802.
- GABUNIA L, VEKUA A, LORDKIPANIDZE D, SWISHER CC, 3RD, FERRING R, JUSTUS A, NIORADZE M, TVALCHRELIDZE M, ANTON SC, BOSINSKI G, JORIS O, LUMLEY MA, MAJSURADZE G & MOUSKHELISHVILI A (2000)
- Earliest Pleistocene hominid cranial remains from Dmanisi, Republic of Georgia: taxonomy, geological setting, and age. Science 288, 1019-1025.
- GIBBONS A (2001)
- The riddle of coexistence. Science 291, 1725-1729.
- GIBBONS A (2002)
- Paleoanthropology. African skull points to one human ancestor. Science 295, 2192-2193.
- GIBERT J, CAMPILLO D, ARQUES JM, GARCIA-OLIVARES E, BORJA C & LOWENSTEIN J (1998)
- Hominid status of the Orce cranial fragment reassessed. J. Hum. Evol. 34, 203-217.
- GUTIN J (1995)
- Remains in Spain now reign as oldest Europeans. Science 269, 754-755.
- HAGAI R & LEVI S (2001)
- When did hominids first leave Africa?: New high-resolution magnetostratigraphy from the Erk-el-Ahmar Formation, Israel. Geology 29, 887-890.
- HARTWIG-SCHERER S (2000)
- Ein möglicher Neandertal-Hybrid und seine Folgen. Stud. Int. J. 7, 84-88.
- HARTWIG-SCHERER S (2002)
- Übergangsform zwischen „Homo“ habilis und Homo erectus? Stud. Int. J. 9, 89-91.
- HAWKS J & WOLPOFF MH (2001)
- Brief communication: Paleoanthropology and the population genetics of ancient genes. Am J Phys Anthropol 114, 269-272.
- INGMAN M, KAESSMANN H, PÄÄBO S & GYLLENSTEIN U (2000)
- Mitochondrial genome variation and the origin of modern humans. Nature 408, 708-713.
- KRINGS M, GEISERT H, SCHMITZ RW, KRAINITZKI H & PÄÄBO S (1999)
- DNA sequence of the mitochondrial hypervariable region II from the neandertal type specimen. Proc. Natl. Acad. Sci. U S A 96, 5581-5585.
- KRINGS M, STONE A, SCHMITZ RW, KRAINITZKI H, STONEKING M & PÄÄBO S (1997)
- Neandertal DNA sequences and the origin of modern humans. Cell 90, 19-30.
- LOEWE L & SCHERER S (1997)
- Mitochondrial Eve: the plot thickens. Trends Ecol. Evol. 12, 422-423.
- MARTINEZ NAVARRO B, TURQ A, BALLESTER JA & OMS O (1997)
- Fuente Nueva-3 (Orce, Granada, Spain) and the first human occupation of Europe. J. Hum. Evol. 33, 611-620.
- MOYA-SOLA S & KOHLER M (1997)
- The Orce skull: anatomy of a mistake. J. Hum. Evol. 33, 91-97.
- OVCHINNIKOV IV, GOTHERSTROM A, ROMANOVA GP, KHARITONOV VM, LIDEN K & GOODWIN W (2000)
- Molecular analysis of Neanderthal DNA from the northern Caucasus. Nature 404, 490-493.
- RELETHFORD JH (2001)
- Ancient DNA and the origin of modern humans. Proc. Natl. Acad. Sci. U S A 98, 390-391.
- REPENNING CA & FEJFAR O (1982)
- Evidence for earlier date of ‘Ubeidiya, Israel, hominid site. Nature 299, 344-347.
- RIGHTMIRE GP (1997)
- Deep roots for the Neanderthals. Nature 389, 917-918.
- ROSAS A & BERMUDEZ DE CASTRO JM (1998)
- On the taxonomic affinities of the Dmanisi mandible (Georgia). Am. J. Phys. Anthropol. 107, 145-162.
- ROSAS A & BERMUDEZ DE CASTRO JM (1999)
- The ATD6-5 mandibular specimen from Gran Dolina (Atapuerca, Spain). Morphological study and phylogenetic implications. J. Hum. Evol. 37, 567-590.
- SCHRENK F (2000)
- Europa von den Hominiden in Schüben besiedelt. FAZ 12. 7. 2000.
- SMITH FH, TRINKAUS E, PETTITT PB, KARAVANIC I & PAUNOVIC M (1999)
- Direct radiocarbon dates for Vindija G(1) and Velika Pecina late Pleistocene hominid remains. Proc. Natl. Acad. Sci. U S A 96, 12281-12286.
- SWISHER CC, CURTIS GH, JACOB T, GETTY AG, SUPRIJO A & WIDIASMORO (1994)
- Age of the earliest known hominids in Java, Indonesia. Science 263, 1118-1121.
- TEMPLETON A (2002)
- Out of Africa again and again. Nature 416, 45-51.
- THORNE AG & WOLPOFF MH (1992)
- Multiregionaler Ursprung der modernen Menschen. Spektrum d. Wiss. Juni, 72-79.
- VLCEK E, MANIA D & MANIA U (2000)
- A new find of a Middle Pleistocene mandible from Bilzingsleben, Germany. Naturwissenschaften 87, 264-265.
- WADDLE D (1994)
- Matrix correlation tests support a single origin for modern humans. Nature 368, 452-455.
|
|  |
