 |
 Studium Integrale Journal
Studium Integrale Journal
20. Jahrgang / Heft 2 - Oktober 2013
Seite 116 - 123
|
|
 |
| Abb. 1: Spinnennetze sind immer wieder für Überraschungen gut. (Foto: W. Borlinghaus) |
|
Verschiedene Eigenschaften von Spinnenseiden, die Webspinnen (Araneae) zum Bau ihrer Netze verwenden, tragen dazu bei, erfolgreich Beute zu fangen. Die Mikrostruktur der Spinnfäden, die Beschaffenheit ihrer Oberflächen, die Konstruktion des Netzes (möglicherweise auch dessen Verzierungen), flüchtige (Geruchs-)Stoffe und viele weitere Details sind daran beteiligt.
Es ist schon seit langem bekannt, dass Insekten sich elektrostatisch aufladen, wenn sie über elektrisch geladene Oberflächen laufen oder im Flug auf geladene Teilchen stoßen. Bei Bienen und Hummeln sind auch Auswirkungen der elektrostatischen Aufladung auf ihren Erfolg beim Pollensammeln untersucht worden.
Über einen Einfluss von elektrischer Ladung beim Beutefang in Netzen von Webspinnen ist bereits spekuliert worden. Jetzt haben Ortega-Jimenez & Dudle (2013) erstmals experimentelle Laboruntersuchungen vorgelegt. Sie haben dazu frisch gefangene und getötete Insekten (Honigbienen [Apis mellifera], Goldfliegen [Lucilia sericata], Fruchtfliegen [Drosophila melanogaster] und Blattläuse [Aphidoidea]) sowie Wassertropfen mit Hilfe eines Generators aufgeladen und in Kontakt mit Netzen der Kreuzspinne (Araneus diadematus) gebracht. Die auf das Netz oder nahe am Netz vorbei fallenden geladenen Objekte sowie die Fäden des Netzes wurden dabei mit einer Videokamera (1500 Bilder/s-1) aufgenommen. Im Versuchsaufbau im Labor konnten deutliche Deformationen der Spinnfäden dokumentiert werden, wenn geladene Insekten oder Wassertropfen sich in deren Nähe bewegten. Die hochelastischen Spiral- oder Fangfäden wurden dabei stärker ausgelenkt als die festeren Rahmen- und Speichenfäden.
Daten über den Einfluss von elektrostatisch geladenen Objekten auf Spinnennetze unter natürlichen Bedingungen liegen derzeit nicht vor. Die von Ortega-Jimenez & Dudle vorgelegten Untersuchungen zeigen nicht nur das Zusammenwirken unterschiedlichster Effekte, die zum erfolgreichen Beutefang von Webspinnen führen. Der unter Laborbedingungen demonstrierte Effekt steigert auch die Effektivität beim Einfang von Pollen und Sporen mit Spinnennetzen. Auch diese finden sich auf dem Speiseplan von Spinnen.
[Ortega-Jimenez VM & Dudle R (2013) Spiderweb deformation induced by electrostatical charged insects. Sci. Reports doi: 10.1038/srep02108]
|

 |
| Abb. 1: Springspinne Hasarius adansoni. (Foto: Nick Hobgood; GNU Freie Dokumentations-Lizenz) |
|
Spinnenseide – ein Biomaterial mit sehr interessanten Eigenschaften – wird von Spinnen für unterschiedliche Anwendungen hergestellt. Radnetzspinnen bauen daraus ihren namengebenden Lebensraum, viele Spinnen nutzen Spinnenseide als Sicherungsfaden, zur schützenden Umhüllung ihrer Gelege, zur Fortbewegung durch die Luft (Altweibersommer, Balooning), u. v. a. m.
Springspinnen befestigen in der Regel vor einem Sprung auf eine Beute einen Seidenfaden auf dem Untergrund. Dieser Faden wurde bisher üblicherweise als Sicherungsfaden interpretiert, der verhindert, dass die Spinne durch das Beutetier verschleppt wird, oder es ermöglicht, dass die Spinne bei einem Fehlversuch wieder an den Ausgangsort zurückkehren kann.
Chen et al. (2013) konnten jetzt in Laboruntersuchungen an bestimmten Springspinnen (Hasarius adansoni) zeigen, dass der an der Absprungstelle fixierte Seidenfaden es der Spinne erlaubt, ihren Körper im Verlauf der Flugphase zu stabilisieren. Mit Hochgeschwindigkeitskameras haben die Autoren im Labor den Sprung von Spinnen aufgenommen und analysiert. Von den für die Untersuchung im Freiland gesammelten Spinnen verwendeten wenige keinen Sicherungsfaden beim Sprung, zeigten darüber hinaus aber keine erkennbaren Besonderheiten; diese dienten zum Vergleich. Bei Springspinnen, die einen Sicherungsfaden an die Absprungstelle heften, weist der Körper während der Sprungphase in der räumlichen Orientierung eine deutlich geringere Variation im Vergleich zu den Exemplaren auf, die keinen Faden verwenden. Die Landung erfolgt ohne Faden unkontrollierter und dauert deutlich länger (keine Bremswirkung). Wenn H. adansoni vor dem Sprung einen Sicherungsfaden an der Absprungstelle anheftet, dann kann sie durch Veränderung des Winkels zwischen Faden und ihrem Hinterleib (Abdomen) die Körperrotation während des Sprungs beeinflussen und bei der Landung bremsen. Dies vergrößert die Erfolgsaussichten bei ihrer Jagd nach Beute.
Chen et al. (2013) schließen ihre Veröffentlichung mit der Bemerkung ab, dass sie mit der beschriebenen Funktion des Spinnfadens neben den aerodynamischen Kräften des Flügelschlags und der Trägheit schwingender Köperanhänge eine neue dritte Einflussgröße zur Kontrolle der Flugstabilisierung gefunden haben. Sie hoffen, diese Entdeckung zukünftig bei der Entwicklung von manövrierbaren Robotern nutzen zu können.
[Chen Y-K, Liao C-P, Tsai F-Y & Chi K-C (2013) More than a safety line: jump-stabilizing silk of salticids. J. R. Soc. Interface 10:20130572; http://rsif.royalsocietypublishing.org/content/10/87/20130572.]
|
|
Die ungefähr 8.000 Arten umfassenden Schwämme (Porifera) sind sehr einfach gebaute Tiere. Sie besitzen weder ein Atmungs- noch ein Muskel-, ein Nerven- oder Ausscheidungssystem. Auch Sinneszellen sind unbekannt. Jede einzelne Zelle sorgt sozusagen hinsichtlich Ernährung, Stoffwechsel und Ausscheidung weitgehend für sich selbst.
Es ist daher naheliegend, die Schwämme in der Systematik an die Basis der vielzelligen Tiere (Metazoa) zu stellen. Diese Position machen ihnen nur die eigenartigen winzigen Placozoa („Plattentiere“ mit der einzigen Art Trichoplax adhaerens) streitig. Nach morphologischen Kriterien stehen die Placozoa an der Basis der Vielzeller, nach molekularbiologischen dagegen die Schwämme. Vor einigen Jahren brachte eine umfangreiche vergleichende Analyse von etwa 9400 genomischen, mitochondrialen, RNA-Struktur- und morphologischen Merkmalen eine Überraschung: Unter evolutionstheoretischen Voraussetzungen mussten die Placozoa zusammen mit den Schwämmen, Nesseltieren (Cnidaria) und Rippenquallen (Ctenophora) von dem Ast abgetrennt werden, der zu den Bilateria führt (zweiseitig symmetrische Tiere, der Großteil aller Tiere) (Schierwater et al. 2009, Blackstone 2009). Die beiden Gruppen gelangten so auf verschiedene Entwicklungsäste innerhalb der Vielzeller und konnten nicht mehr in eine Evolutionsabfolge gestellt werden.
Die stammesgeschichtliche Position der basalen Vielzeller war zuvor schon sehr kontrovers diskutiert worden, da es viele einander widersprechende Datensätze gibt (Schierwater et al. 2009, vgl. dortige Fig. 1). Die Verwandtschaftsverhältnisse scheinen zunehmend verwirrender zu werden. Denn aufgrund neuer Genom-Sequenzdaten der Rippenqualle Pleurobrachia bachei sind die Rippenquallen überraschenderweise tiefer im evolutionstheoretischen Stammbaum als die Schwämme zu stellen (Moroz et al. 2013; vgl. Maxmen 2013). Dies steht in klarem Widerspruch zu den morphologischen Daten; schließlich sind die Rippenquallen erheblich komplexer gebaut. Sie besitzen Nerven, Muskeln, Gewebeschichten und Lichtsinnesorgane – mit all dem können die Schwämme wie erwähnt nicht aufwarten. Bislang war angenommen worden, dass die Rippenquallen sehr viel später als die Schwämme entstanden sind. Die neuen Daten widersprechen dieser Sicht. Nach einer weiteren Studie an der Rippenqualle Mnemiopsis sind die Rippenquallen entweder ebenfalls basaler als die Schwämme einzuordnen oder mit diesem zusammen an die Basis der Vielzeller zu stellen (Maxwell et al. 2013; vgl. Maxmen 2013). Auch Genfamilien, Zell-Signal-Netzwerke und Genexpressionsmuster der Rippenquallen unterstützen deren unerwartet frühen Ursprung. So fehlt den Rippenquallen z. B. microRNA, das sind kurze RNA-Stücke, die bei fast allen Tiergruppen an der Genregulation beteiligt sind, was als Indiz für „Ursprünglichkeit“ gewertet wird.
Eine der Folgen der Umgruppierung in der Systematik der Tierstämme ist, dass eine zweimalige unabhängige Entstehung des Nervensystems angenommen werden muss (vgl. Pennisi 2013) – eine ebenfalls unerwartete Konsequenz. Trotz erheblicher Ähnlichkeiten mit Nervensystemen anderer Tiere sind dessen genetische Grundlagen bei den Rippenquallen jedoch deutlich verschieden (Maxmen 2013).
Außerdem muss nun angenommen werden, dass die Schwämme durch Verlust von zuvor vorhandenen Eigenschaften entstanden sind. Beide Konsequenzen sind evolutionstheoretisch problematisch. Andererseits sind Schwämme physiologisch nicht primitiv. Insgesamt sind sie demnach eine klar abgrenzbare Gruppe. Einmal mehr zeigt sich, dass die Gewinnung neuer Daten nicht automatisch zunehmende Klarheit über Verwandtschaftsverhältnisse bringt. Durch die hier vorgestellten Erkenntnisse werden diese komplizierter und evolutionäre Erklärungen schwieriger.
[Blackstone NW (2009) A New Look at Some Old Animals. PLoS Biol 7(1), http://dx.doi.org/10.1371/journal.pbio.1000007; Maxmen A (2013) Genome reveals comb jellies’ ancient origin. Nature, doi:10.1038/nature.2013.12176; Maxwell EK, Ryan JF, Schnitzler CE, Browne WE & Baxevanis AD (2012) MicroRNAs and essential components of the microRNA processing machinery are not encoded in the genome of the ctenophore Mnemiopsis leidyi. BMC Genomics 2012, 13:714, doi:10.1186/1471-2164-13-714; Moroz LL, Kohn A, Citarella M, Grigorenko A, Kocot K, Halanych K & Rogaev E (2013) The Genome of the Ctenophore Pleurobrachia bachei: Molecular Insights into Independent Origins of Nervous Systems. http://sicb.org/meetings/2012/schedule/abstractdetails.php?id=527; Pennisi E (2013) Nervous System May Have Evolved Twice. Science 339, 391; Schierwater B, Eitel M, Jakob W, Osigus HJ, Hadrys H, Dellaporta SL, Kolokotronis SO, DeSalle R (2009) Concatenated Analysis Sheds Light on Early Metazoan Evolution and Fuels a Modern „Urmetazoon“ Hypothesis. PLoS Biol. 7(1), http://dx.doi.org/10.1371/journal.pbio.1000020]
|
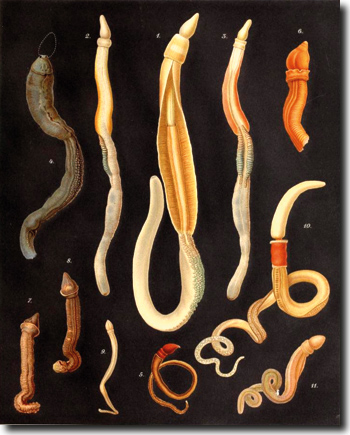 |
| Abb. 1: Bunte Vielfalt von Eichelwürmern. (Aus Spengel, J. W.: Die Enteropneusten des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Berlin 1893) |
|
Die größte Diskontinuität in der Fossilgeschichte ist zweifellos das plötzliche fossile Erscheinen verschiedenster Tiergruppen im geologischen System des Kambriums (Valentine 2004, Erwin & Valentine 2013). Darwin sah in diesem Befund eine mögliche Bedrohung für seine gradualistische Evolutionstheorie. Im Laufe der Zeit wurden seither viele weitere Fossilien entdeckt, auch unterhalb der Präkambrium-Kambrium-Grenze, doch nur wenige vorkambrische Formen sind als Vorläufer der kambrischen Vielfalt diskutabel, und viele kambrische Funde haben die Diskontinuität verstärkt.
Eine der bekanntesten Fossillagerstätten, die wesentlich zur Kenntnis der kambrischen Vielfalt beigetragen haben, ist der mittelkambrische Burgess-Schiefer Kanadas (auf ca. 505 Millionen Jahre datiert), dessen „irre Wundertiere“ von Stephen J. Gould (1989) ausführlich einem breiten Publikum vorgestellt werden. Aus dem Burgess-Schiefer sind auch seit über 100 Jahren wurmartige Organismen bekannt, die als Spartobranchus tenuis klassifiziert wurden. Aber erst kürzlich wurde durch eine genaue Analyse (Caron et al. 2013) nachgewiesen, dass es sich dabei eindeutig um Eichelwürmer handelt, eine Tiergruppe, die zusammen mit den Pterobranchia (Flügelkiemer) zu den Hemichordaten („Halbchordatiere“) gehören (s. Kasten; vgl. auch Gee 2013). Die bis zu 10 cm langen Eichelwürmer leben am Meeresboden und graben Gänge. Sie bewegen sich im Meeresboden durch wellenförmige Muskelkontraktionen vorwärts und leben von im Schlamm enthaltenen organischen Partikeln mit Hilfe eines Nahrungstrichters am Vorderende.
|
„Halbe Chordatiere“; auch als Kiemenlochtiere bezeichnet; Tierstamm mit weniger als 100 bekannten Arten; diese besitzen einen weichen, wurmähnlichen, aber innen ungegliederten Körper, sind äußerlich dreigeteilt (Kopfschild, Kragen und Rumpf). Werden traditionell als Urform der Chordatiere angesehen, zu denen auch die Wirbeltiere gehören (die Chorda ist ein flüssigkeitsgefüllter Stützstab); ihre systematische Stellung ist aber umstritten.
|
Erstaunlich ist, dass die kambrischen Eichelwürmer heutigen Formen aus der Gruppe der Harrimaniiden gleichen. „Es ist fast so, als wenn man ein Foto eines heutigen Exemplars machen würde“ wird Christopher Cameron zitiert, der heutige Eichelwürmer untersucht (http://www.cbc.ca/news/technology/story/2013/03/13/science-phallus-fossil-acorn-worm.html). Bislang waren die Eichelwürmer erst aus der Trias bekannt (nach radiometrischer Datierung vor 200-250 Millionen Jahren). Die Eichelwürmer tragen damit nicht nur zur „Verstärkung“ der kambrischen Explosion bei, sondern sind auch ein Beispiel für Stasis – das nahezu unveränderte „Stehenbleiben“ eines Bauplans über geologische Zeiträume hinweg.
Da auch die Pterobranchia (s. o.) aus dem Kambrium bekannt sind, ist klar, dass der Ursprung der Hemichordaten entsprechend ins Unterkambrium verlegt werden muss (Caron et al. 2013, 503). Einen Unterschied gibt es jedoch bei den fossilen Formen im Vergleich zu den heutigen. Bei den Fossilien wurden häufig faserige Röhren gefunden; in einem Fall war ein Eichelwurm mit dieser Röhre assoziiert. Daher kann man annehmen, dass die kambrischen Eichelwürmer anders als die heutigen zeitweise in Röhren lebten. Dieser Befund wurde von S. Conway Morris, einem der Bearbeiter so interpretiert, dass es sich hier um einen Hinweis handle, dass die fossilen Eichelwürmer Übergangsformen zu den Pterobranchia seien (http://www.bbc.co.uk/nature/21745029), die generell solche Röhren bilden. (Deren Röhren können viel leichter fossilisiert werden und sind als Graptolithen bekannt.) Doch warum aufgrund des Besitzes solcher Röhren die kambrischen Eichelwürmer einen evolutionären Übergangsstatus haben sollen, ist unklar. Viel plausibler ist ein Szenario, wonach die Eichelwürmer ursprünglich ebenso wie die Pterobranchia solche Röhren ausgebildet haben, diese später aber verloren haben (Caron et al. 2013, 505).
[Caron JB, Conway Morris S & Cameron CB (2013) Tubicolous enteropneusts from the Cambrian period. Nature 495, 503-506; Erwin DH & Valentine JW (2013) The Cambrian explosion. The construction of animal biodiversity. Greenwood Village, Colorado; Gee H (2013) Tubular worms from the Burgess Shale, Nature 495, 458-459; Gould SJ (1989) Wonderful life. The Burgess shale and the nature of history. New York; Valentine JW (2004) On the origin of phyla. Chicago and London]
|
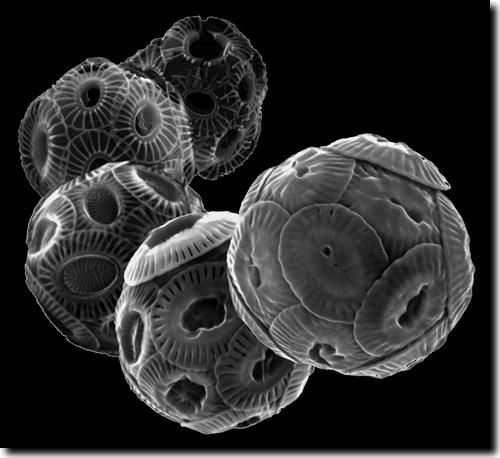 |
| Abb. 1: Morphotypen der Kalkalge Emiliania huxleyi mit unterschiedlichem Kalzifizierungsgrad. Vorne: stark kalzifizierte Organismen, der Kalzifizierungsgrad nimmt nach hinten ab. (Foto: Luc Beaufort, CEREGE, Univ. Aix-Marseille/CNRS) |
|
Während man sich an Meldungen von neu sequenzierten Genomen von immer mehr Lebewesen inzwischen fast gewöhnt hat, ist die Ermittlung des gesamten Erbguts von Mikroorganismen nach wie vor eine große Herausforderung. Das hat u. a. folgenden Grund: Nach derzeitigen Erkenntnissen stimmt die DNA-Sequenz im Erbgut zweier Menschen zu ca. 99 % überein, dagegen beträgt z. B. die Übereinstimmung bei zwei Stämmen der Kalkalgenart Emiliania huxleyi nur 70 oder 80 %. Was also ist die DNA-Sequenz dieser einzelligen Alge? Dieses Phänomen ist typisch für Mikroorganismen und erschwert die Analyse von deren Genom. Bei Bakterien hat man daher das Konzept des Pan-Genoms entwickelt und erfolgreich genutzt. Dabei ermittelt man den Genbestand, der bei allen Vertretern einer Art vorliegt (Kerngenom) und beschreibt dann summarisch die einzelnen Gene, die darüber hinaus bei verschiedenen Stämmen vorkommen; die Summe über all diese Gene wird als Pan-Genom bezeichnet. Das Pan-Genom von E. huxleyi ist jetzt erstmals bestimmt worden. Dafür hat eine 75-köpfige Autorengruppe vierzehn verschiedene Stämme von E. huxleyi sequenziert (Read et al. 2013).
Die Bedeutung der Kalkalgen (Coccolithophorales) liegt darin, dass sie durch Algenblüten in Meeres-Ökosystemen in ihren filigranen Kalkhüllen enorme Mengen an CO2 binden und damit Einfluss auf die Entwicklung klimatischer Verhältnisse nehmen.
Das Genom des E. huxleyi-Stammes CCMP1516 besteht aus 141,7 Millionen Basenpaaren und enthält 30 569 proteincodierende Gene. Etwa 2/3 davon scheinen zum Kerngenom zu gehören; etwa 25% der in CCMP1516 gefundenen Gene lassen sich in mindestens drei der anderen untersuchten Stämme nicht nachweisen. Im Erbgut dieses einzelligen Organismus sind also mehr Proteine codiert als im Genom des Menschen – die Zahl der proteincodierenden Gene korreliert also nicht mit der Komplexität der Organismen.
Im erstaunlich variablen Genom dieser Kalkalgen sehen die Autoren einen Ansatz zur Erklärung der phänotypischen (gestaltlichen) Variabilität sowie der physiologischen und damit verbunden auch ökologischen Vielfalt von E. huxleyi. Möglicherweise eröffnet die hier vorgestellte Studie den Zugang zu weiteren Erkenntnissen, wie das Genom nicht als „Buch des Lebens“, sondern als eine Komponente von Organismen in Wechselwirkung mit anderen Einflussgrößen des Ökosystems Lebensformen prägt.
[Read B, Kegel J, Klute MJ et al. (2013) Pan genome of the phytoplankton Emiliania underpins its global distribution. Nature; doi: 10.1038/nature12221; Pressemitteilung des Alfred Wegener Instituts: http://tinyurl.com/qj6ohxc]
|
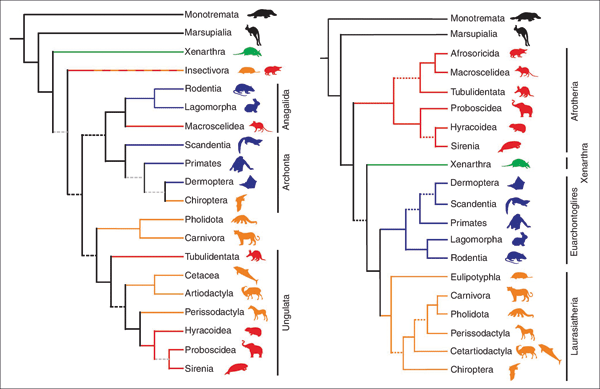 |
Abb. 1: Links der bislang akzeptierte Baum der plazentalen Säugetiere nach morphologischen Merkmalen, rechts der sich abzeichnende Baum nach molekularen Daten. (Aus Springer et al. 2004)
|
|
Die klassische evolutionsbiologische Sicht der Säugetierevolution lässt sich wie folgt zusammenfassen: Während der Zeit der Dinosaurier (Mesozoikum) existierten nur einzelne insektenjagende Vertreter urtümlicher Säugetiergruppen, die sich deutlich von den heutigen unterschieden. Der Einschlag eines großen Asteroiden vor ca. 66 Millionen Jahren schuf u. a. durch die darauf folgende weltweite Umweltzerstörung, durch den Klimawandel und das Aussterben der Dinosaurier den Freiraum für den Siegeszug der modernen plazentabesitzenden Säugetiere (Plazentalier). Innerhalb nur weniger Jahrmillionen, so die Geschichte weiter, traten zahlreiche Vertreter der modernen Ordnungen dieser Säugetiergruppe explosionsartig auf, begleitet von der ebenso rasch aufblühenden Vielfalt unter den Beuteltieren (Marsupialier), die einen anderen modernen Zweig der Säugetiere repräsentieren. Die klassische biologische Systematik der Säugetiere und die Fossilüberlieferung stützte dieses Bild bis zum Ende des 20. Jahrhunderts.
In den letzten zwanzig Jahren haben zahlreiche neue Fossilfunde und ein nahezu unüberschaubarer Zuwachs molekulargenetischer Daten das klassische Bild zur Evolution der Säugetiere jedoch aufgelöst. Tiefgreifende Änderungen bisheriger Modelle zur Systematik und Evolution der Säuger wurden vollzogen, andere sind nur teilweise akzeptiert oder werden unvermindert kontrovers diskutiert (Yoder et al. 2013).
Die bis zur Jahrtausendwende geltende Systematik der plazentalen Säugetiere unterschied auf der Grundlage morphologischer Kriterien die basale Überordnung die Xenarthra („Nebengelenktiere“ wie Ameisenbär, Faultier und Gürteltiere) von den Überordnungen der Ungulata (Huftiere, u. a. mit Seekühen, Rüsseltieren, Unpaarhufern, Paarhufern), der Archonta (u. a. mit Primaten, Fledermäusen, Spitzhörnchen) und der Anagalida (wie Nagetiere, Hasenartige, Spitzmaus) (Abb. 1, links). Die Integration von molekularen Daten änderte diese Gruppierung tiefgreifend. Der auf molekulargenetischen Daten basierende Stammbaum, den Springer et al. (2004) vorstellten, ordnet die 18 plazentalen Säugetierordnungen ebenfalls vier Überordnungen zu: Xenarthra, Afrotheria, Laurasiatheria und Euarchontoglires. Die zuletzt genannten drei neu geschaffenen Überordnungen tragen ihren Namen aufgrund der Region, in der ihre stammesgeschichtliche Abspaltung von den übrigen Überordnungen mit den ihnen zugeordneten Säugetierordnungen und -familien vermutet wird. Diese neuen Überordnungen fassen z. T. Ordnungen plazentaler Säugetiere auf eine Weise zusammen, wie sie nach morphologischen Kriterien nie zusammengestellt würden (Abb. 1, rechts). Z. B. stehen die Fledermäuse nun den Huftieren näher als den Primaten, und die Elefantenspitzmäuse (Rüsselspringer) gehören nicht mehr zu den Insektenfressern oder Hasenartigen, sondern sind eine Untergruppe der Afrotheria und damit enger verwandt mit den Erdferkeln.
 |
| Abb. 2: Madagassischer (links) und europäischer Igel. Eines von vielen Konvergenzbeispielen unter Säugetieren. (GNU Freie Dokumentations-Lizenz) |
|
Nach Bininda Emonds et al. (2012) haben die vergleichend systematischen Forschungen der letzten Jahre dennoch eine weitestgehende Übereinstimmung von morphologischen und molekularbiologischen Datensätzen innerhalb der Xenarthra, Laurasiatheria und Euarchontoglires gezeigt (allerdings nur um den „Preis“ der Annahme vieler Konvergenzen, wenn diese Systematik stammesgeschichtlich interpretiert wird, s. u.). Bei den Afrotheria ist dies jedoch nicht gelungen. Der stammesgeschichtliche Ursprung dieser äußerlich sehr inhomogen erscheinenden Gruppe, zu der z. B. Seekühe, Erdferkel, Elefanten und die Goldmulle zählen, wird in Afrika vermutet.
Aufgrund der auch durch jüngere DNA-Sequenzanalysen gefestigten, modernen Systematik muss davon ausgegangen werden, dass im Rahmen der evolutionären Diversifikation der Säugetiere innerhalb und zwischen den einzelnen Überordnungen zahlreiche und ausgeprägte konvergente Merkmalsentwicklungen erfolgt sein müssen (z. B. Konvergenz zwischen madagassischem und europäischem Igel, Abb. 2). Die extrem hohe Anzahl von Konvergenzen, die bei den plazentalen Säugetieren, ob rezent oder fossil) anzunehmen ist, bezeichneten Springer et al. als Phänomen der korrelierenden Merkmalsevolution. Springer et al. (2004) forderten deshalb, dass künftige phylogenetische Modellierungen das Phänomen der korrelierenden Merkmalsevolution berücksichtigen müssen. Eine befriedigende evolutionstheoretische Erklärung dieses Phänomens, das laufend durch neue Befunde bestätigt wird, steht immer noch aus (vgl. dazu auch Ullrich 2012).
[Bininda-Emonds ORP et al. (2007) The delayed rise of present-day mammals. Nature 446, 507-512; Bininda-Emonds ORP et al. (2012) Rocking clocks and clocking rocks: a critical look at divergence time estimation in mammals. In: Asher JR & Müller J (eds) From Clone to Bone - The Synergy of Morphological and Molecular Tools in Palaeobiology. Cambridge, pp. 38-82; Yoder AD et al. (2013) Fossils versus Clocks. Science 339, 656-658; O’Leary MA et al. (2013) The Placental Mammal Ancestor and the post-K-Pg Radiation of Placentals. Science 339, 662-667; Springer MS et al. (2004) Molecules consolidate the placental mammal tree. Trends Ecol. Evol. 19, 430-438; Ullrich H (2012) Verwirrende Konvergenzen. Zur Evolution des Mittelohres der Säugetiere. Stud. Int. J. 19, 20-33.]
|
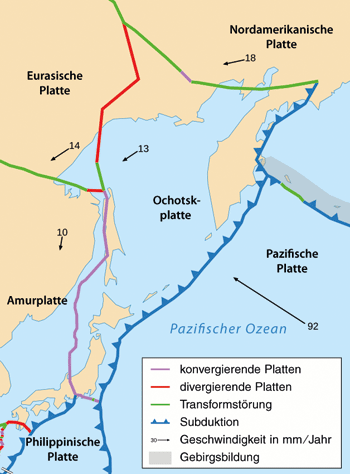 |
| Abb. 1: Die Lage sowie aktuelle Bewegungsrichtung und -geschwindigkeit der Erdplatten vor Japan. (Grafik: MARUM, Universität Bremen; freundliche Überlassung durch MARUM) |
|
PDF-Version des Streiflichts
Das Seebeben von Tohoku mit einer Magnitude von 9,0 löste eine Kettenreaktion aus: Ein bis zu 15 m hoher Tsunami verwüstete im Nordosten Japans ganze Küstenabschnitte und setzte das küstennahe Kernkraftwerk Fukushima-Daiichi unter Wasser, der Beginn eines nicht mehr abzuwendenden Super-GAUs.
Die geowissenschaftliche Analyse der Ereignisse, die dem Seebeben gefolgt sind, ist noch nicht abgeschlossen. Kürzlich berichteten Strasser et al. (2013) in der August-Ausgabe von Geology, dass aufgrund des Tohoku-Bebens im Japan-Graben ein großflächiges Sedimentpaket absackte und sich die Grenzen der beteiligten Erdplatten um zwei bis drei Kilometer verschoben. Dazu Gerold Wefer, Co-Autor und damaliger MARUM-Direktor, zitiert in MARUM.de: „Als Geowissenschaftler sind wir es gewohnt, in sehr langen Zeiträumen von Jahrhunderttausenden oder gar -millionen zu denken.“ Und weiter: „Diese Expeditionen haben uns gezeigt, dass Plattengrenzen bisweilen auch von plötzlichen Ereignissen in Sekunden oder Minuten tiefgreifend verändert werden können.“
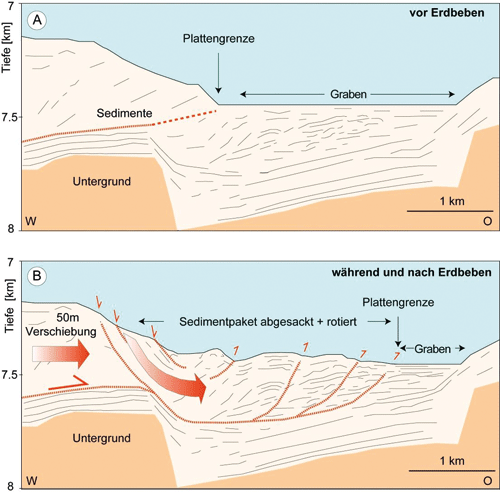 |
| Abb. 2: Der Japan-Graben vor und nach dem Tohoku-Erdbeben am 11. März 2011. (Grafik: M. Kölling, MARUM, Universität Bremen, nach: Strasser et al. in Geology, August 2013; freundliche Überlassung durch MARUM) |
|
Der Artikel präsentiert Ergebnisse von Untersuchungen mit den Forschungsschiffen Mirai (Japan) und Sonne (MARUM, Deutschland) im Jahre 2012, die 140 km östlich des Epizentrums, unmittelbar am Abhang des bis zu 7,6 km tiefen Japan-Grabens vorgenommen wurden. Dort befindet sich die Meeresboden-Nahtlinie, an welcher die Pazifische Platte unter die Ochotsk-Platte mit dem japanischen Vulkanbogen abtaucht (Abb. 1). Die Studie basiert auf einer hochauflösenden Echolotkartierung des Meeresbodens entlang einer Profillinie senkrecht zum Grabenverlauf und einem Vergleich mit Daten aus 1999 und 2004. Darüber hinaus sind entlang dieser Profillinie mehrere bis zu 9 m lange Sedimentkerne gezogen und ausgewertet worden.
Eine besondere Herausforderung war, die beobachteten Veränderungen des Meeresbodens einem (bzw. dem Tohoku-) Erdbeben eindeutig zuordnen zu können. Ein erster Datierungsansatz erfolgte mit der 210Pb-Methode (Halbwertszeit des kurzlebigen radioaktiven 210Pb: 22,3 Jahre), welche das Alter auf weniger als das Fünffache der Halbwertszeit, also < 110 Jahre eingrenzte. Ein anderer Ansatz bestand darin, für zwei der Sedimentkerne geochemische Porenwasser-Profile (mit der Tiefe) aufzustellen und insbesondere die SO42-Konzentrationen zu betrachten. Diese wiesen im oberen Bereich der Sedimentsäule einen knickförmigen Verlauf auf, der auf eine gravierende Porenwasserstörung hinwies. Nach Modellberechnungen würde sich ein Porenwasser-Gleichgewicht nach etwa 10 Jahren einstellen; der jeweilige knickförmige Verlauf zeichnete demnach einen Zeitpunkt von etwa 0,5 – 2 Jahren nach dem Störungsereignis nach. Die zwei Kerne waren im März 2012, also 1 Jahr nach dem Ereignis gezogen worden. Folglich ist die Wahrscheinlichkeit sehr groß, dass die Deformationsstrukturen vom Tohoku-Beben ausgelöst worden waren.
Im Ergebnis gehen die Autoren von folgendem Szenario aus (Abb. 2): Am Rand des Japan-Grabens löste das Erdbeben einen ruckartigen Versatz der Erdkruste um 50 m aus. Dabei sackten großflächig (etwa 28 km2) Sedimentpakete ruckartig als zusammenhängende Blöcke in die Tiefe und stauchten an manchen Stellen den Meeresboden. Die so entstandene unruhige Bodenoberfläche aus Wülsten und Trögen liegt bis zu 50 m höher als vor dem Erdbeben. Der Meeresboden am Hang dagegen liegt nunmehr etwa 50 m tiefer. Die oberflächennahe Grenze der Erdplatten wurde um mehr als 2 km nach Osten versetzt: „(...) within a single event during a period of seconds or minutes.“
[Strasser M, Kölling M, Santos Ferreira C dos, Fink HG, Fujiwara T, Henkel S, Ikehara K, Kanamatsu T, Kawamura K, Kodaira S, Römer M, Wefer G & the R/V Sonne Cruise SO219A & JAMSTEC Cruise MR12-E01 scientists (2013) A slump in the trench: Tracking the impact of the 2011 Tohoku-Oki earthquake. Geology 41, 935-938; http://www.marum.de/Turbulenzen_in_der_Tiefe.html, Abfrage 19. 08. 2013; http://earthquake.usgs.gov/earthquakes/eqinthenews/2011/usc0001xgp/, Abfrage 19. 08. 2013]
|
|
 |
| Abb. 1: Große Wachsmotte Galleria mellonella. (GNU Freie Dokumentations-Lizenz) |
|
Das „evolutionäre Wettrüsten“ zwischen nachtaktiven Motten und Fledermäusen gehört zu den am besten erforschten Räuber-Beute Beziehungen in der Biologie. Die Echoortung der Fledermäuse, das sie zum Navigieren und Jagen benutzen, ist allgemein bekannt. Fast alle Fledermausarten der Alten Welt verfügen über diese Fähigkeit. In der Öffentlichkeit weniger bekannt ist die Fähigkeit mehrerer Mottenspezies, die von den Fledermäusen ausgestoßenen Ultraschalltöne zu hören. So ist die Beute frühzeitig gewarnt, wenn ein Jäger im Anflug ist, und kann sich in Sicherheit bringen.
Manche Fledermäuse haben sich daran angepasst, indem sie beispielsweise Frequenzen benutzen, die zu hoch oder zu tief sind, um von Motten gehört zu werden. Die Motten passen sich ihrerseits mancherorts an diese veränderte Frequenz an; so ist z. B. die optimale Hörfrequenz von Motten auf Hawaii tiefer als sonst, passt aber zu der Frequenz, die die einzige dort einheimische Fledermausart verwendet. Auch andere Verhaltensänderungen wurden schon auf „beiden Seiten“ beobachtet. Diese Situation kann als ein evolutionäres Wettrüsten interpretiert werden, bei dem sich die Kontrahenten immer wieder anpassen, um einen Vorteil gegenüber dem anderen zu erringen.
Bisher war die höchste Frequenz, von der man wusste, dass eine Mottenart sie hören kann, 150 kHz beim in Nordamerika lebenden Schwammspinner Lymantria dispar. (Zum Vergleich: Menschen hören Töne maximal zwischen 20 Hz und 20 kHz.) Normalerweise liegt die optimale Hörfrequenz bei nachtaktiven Motten (Noktuiden) bei 20-50 kHz. Die höchst-bekannte, von Fledermäusen emittierte Frequenz liegt jedoch bei 212 kHz und ist damit signifikant höher. Deshalb fragte sich ein britisches Forscherteam, ob es Motten gibt, die diese Frequenz auch noch hören können, die also im Wettrüsten mit den Fledermäusen mithalten (Moir et al. 2013). Als Untersuchungsobjekt wählten sie die Große Wachsmotte Galleria mellonella, die weltweit verbreitet ist und daher mit einer Vielzahl von Fledermausarten in Kontakt kommt (Abb. 1). Sie testeten die Motten auf Sensibilität für Frequenzen zwischen 50-300 kHz. Das erstaunliche Ergebnis: Selbst die höchste Frequenz von 300 kHz wurden von den Ohren der Motten registriert! Damit können diese Motten höhere Töne hören als jedes andere bekannte Tier. Das ist umso erstaunlicher, wenn man bedenkt, dass die Ohren von Motten aus höchstens vier Gehörzellen (wie bei G. mellonella) bestehen, die überdies noch nicht einmal in der Lage sind, zwischen Frequenzen zu unterscheiden, sondern nur entweder stimuliert sind oder nicht. Aus diesem Grund benutzen die Autoren auch vielsagende Anführungszeichen im Titel ihres Artikels: „Extrem hohe Frequenz-Sensibilität in einem ‚einfachen‘ Ohr“.
Dieses Resultat wirft aber gleichzeitig neue Fragen auf. Wie oben erwähnt ist keine Fledermaus bekannt, die solch hohe Töne verwendet. Es könnte natürlich sein, dass man einfach noch nicht lange genug geforscht hat, oder dass man aufgrund technischer Schwierigkeiten bei früheren Messungen diese Frequenzen übergangen hat. Dies ist aus physikalischen Gründen jedoch unwahrscheinlich: Berechnungen zufolge ist die atmosphärische Schalldämpfung ab etwa 200 kHz zu groß, um eine effektive Echoortung zu ermöglichen. Deshalb ist nicht zu erwarten, dass irgendwelche Fledermäuse Ultraschall mit diesen hohen Frequenzen benutzen. Aber warum sollten Motten dann Töne hören können, die kein Jäger einsetzt? Die Autoren schlagen vor, dass die Sensibilität für diese Frequenzen nur ein Nebeneffekt der eigentlich vorteilhaften Anpassung sind: Gehörzellen, die auf hohe Frequenzen ausgelegt sind, können wegen ihrer Mechanik bei niedrigeren Frequenzen schneller reagieren. So kann G. mellonella schneller auf die Frequenzen reagieren, denen sie tatsächlich ausgesetzt ist, ein Effekt, der sich im Labor bestätigen ließ.
Kritisch zu bewerten ist der Eindruck, den Moir et al. in der Kurzzusammenfassung ihres Artikels entstehen lassen. Sie schreiben: „Mit einer im Tierreich bisher unerreichten Tonfrequenz-Sensibilität ist die Große Wachsmotte vorbereitet und bewaffnet für jegliche Anpassung am Echolot der Fledermaus, die diese im Zuge des anhaltenden evolutionären Krieges zwischen Fledermaus und Motte vornehmen mag“ (Moir et al. 2013, übersetzt vom Autor). Dies suggeriert, dass Anpassungen im Voraus entstehen können, was evolutionstheoretisch gesehen sehr problematisch ist. Schließlich kann die natürliche Selektion nur auf Umstände reagieren, die schon vorhanden sind; sie ist, um es mit Dawkins zu sagen, ein „blinder Uhrmacher“. Eine Voranpassung dieser Art würde in der Tat besser zum Modell der programmierten Variabilität passen, in der eine polyvalente Stammform schon alle nötigen Allele besitzt oder durch wenige Mutationen erhalten kann, die sie einmal brauchen wird.
[Moir HM, Jackson JC & Windmill JFC (2013) Extremely high frequency sensitivity in a ‘simple’ ear. Biol. Lett. 9, 20130241; Waters DA (2003) Bats and Moths: What is there left to learn? Physiol. Entomol. 28, 237-250; Conner WE & Corcoran AJ (2012) Sound strategies: The 65-million-year-old battle between bats and insects. Ann. Rev. Entomol. 57, 21-39.]
|
|
 |
| Abb. 1: Blütenstand der Knoblauchsrauke Alliaria petiolata. (Foto: R. Junker) |
|
Bioarchäologen haben bildlich gesprochen in prähistorische Kochtöpfe geschaut und dabei klare Hinweise auf die Verwendung von Samen der Knoblauchsrauke (Alliaria petiolata, Abb. 1) gefunden. Die Tonscherben der Gefäße waren an der deutschen Ostseeküste in Neustadt an der Lübecker Bucht und in Dänemark gefunden worden. Die Datierung der Proben ergab ein Alter von 5460 ± 90 bis 5122 ± 63 Jahre vor der Gegenwart (6450 bis 5700 cal BP[kalibriert]). Dazu waren verkohlte Probenbestandteile und mit den Tonscherben assoziierte Säugetierknochen mittels Radiokarbon-Untersuchungen (14C) datiert worden.
Mikroskopisch konnten Saul et al. (2013) die kugelig aufgewölbten Phytolithen (mineralisierte – meist verkieselte – pflanzliche Bestandteile, die gesteinsbildend sein können) durch Vergleich mit heutigen Phytolithen aus dem Samengewebe der Knoblauchsrauke identifizieren. Die Autoren sehen darin einen klaren Hinweis auf die Verwendung der Knoblauchsrauke als Gewürz. Die Blätter der Knoblauchsrauke bringen im Salat ein knoblauchartiges Aroma und die Samen können aufgrund ihres Gehalts an Senföl-Verbindungen (wie z. B. das Sinigrin) als Gewürz verwendet werden (Düll & Kutzelnigg 2011). Aufgrund von Lipidanalysen vermuten Saul et al., dass Fleisch von Meerestieren und Wiederkäuern zusammen mit pflanzlichen Zutaten die eigentlichen Nahrungsmittel waren.
Der Zeitraum, in den die untersuchten Funde eingeordnet werden (spätes Meso- und Neolithikum), entspricht nach gängigen Vorstellungen im westlichen Baltikum dem Übergang von Jäger- und Sammlerkulturen zur Landwirtschaft. Egal ob die dort lebenden Menschen die Knoblauchsrauke aus eigenem Antrieb zum Würzen ihrer Speisen nutzten oder ob sie eine solche Tradition aus dem Nahen Osten übernommen haben, Saul et al. belegen jedenfalls, dass Menschen sehr früh begonnen haben, natürliche Pflanzen nicht nur als Nährstoffe, sondern auch gezielt zur Beeinflussung des Geschmacks zu verwenden.
[Doran PT, Fritsen CH, Murray AE, Kenig F, McKay CP & Kyne JD (2008) Entry approach into pristine ice sealed – Lake Vida, East Antarctica, a model ecosystem. Limnol. Oceanogr. Methods 6, 542-547; Murray AE, Kenig F, Fritsen CH, McKay CP, Cawley KM, Edwards R, Kuhn E, McKnight DM, Ostrom NE, Peng V, Ponce A, Priscue JC, Samarkin V, Townsend AT, Wagh P, Young SA & Doran PT (2012) Microbial life at -13 °C in the brine of an ice-sealed Antarctic lake. Proc. Natl. Acad. Sci. USA, doi: 10.1073/pnas.1208607109]
|
|
|
 |
