 |
Molekulare Daten belegen: Konvergente Evolution
komplexer Merkmalssysteme bei den Singvögeln
Neue Ergebnisse und alternative Deutung aus einer teleologischen Perspektive
von Christoph Heilig
Studium Integrale Journal
16. Jahrgang / Heft 1 - Mai 2009
Seite 45 - 48
|
 |
Zusammenfassung: Die Singvogel-Familie der Honigfresser (Meliphagidae) beinhaltet ca. 180 Arten, welche man vor allem im australasischen Raum antreffen kann. Sie waren aber auch durch fünf endemische Arten (in zwei Gattungen) auf Hawaii beheimatet, die mittlerweile jedoch als ausgestorben gelten. Ihre Zugehörigkeit zu den Meliphagidae, mit deren übrigen Vertretern sie eine Vielzahl von Anpassungen an die Ernährung durch Pollen und Nektar teilen, wurde bisher nie in Frage gestellt. Ohne die nun erfolgte Analyse molekularer Merkmale auf der Basis von Museumsexemplaren wäre das wohl auch nie geschehen. Diese belegen jedoch, dass die monophyletische Gruppe (= Gruppe mit gemeinsamem Vorfahren) der heute ausgestorbenen Hawaiianischen Gattungen Moho und Chaetoptila nicht den Honigfressern zuzurechnen sind. Tatsächlich ergaben die Sequenz-Analysen sogar eine sehr entfernte Position der beiden Gattungen zur Familie der Meliphagidae. Demnach sind sie stattdessen inmitten von morphologisch unähnlichen Insekten- und Fruchtfressern anzusiedeln.
Aufgrund dieser Ergebnisse führen die Autoren der Studie die neue Familie Mohoidae für die beiden Gattungen ein. Damit liegt nun erstmals eine Vogelfamilie vor, welche in den letzten zweihundert Jahren komplett ausstarb. Aus evolutionsbiologischer Sicht ist bedeutsam, dass die Anpassungen der Mohoidae an den Nektarkonsum konvergent zu den vergleichbaren Merkmalen der Meliphagidae entstanden sein sollen. Da diese Konvergenzen auch relativ nahrungsunspezifische Bereiche wie das Federkleid und den Gesang betreffen, ist unklar, wie entsprechende parallele Selektionsdrücke aussehen sollten.
Ausgehend von einem gemeinsamen Anpassungsreservoir an verschiedene Nahrungsangebote in den Stammformen der beiden Familien lassen sich die Daten jedoch problemlos deuten. Die Hypothese von gemeinsamem Potential – entweder in einem gemeinsamen Vorfahren oder im Sinne eines parallelen Designs – erweist sich einmal mehr als sinnvoller Forschungsansatz für die teleologische Perspektive und eröffnet die Möglichkeit einer progressiven Entwicklung des eigenen Forschungsprogramms (im Sinn von Lakatos 1974) gegenüber der herkömmlichen – ateleologischen – Deutung des Lebens. Ein genetisches Potential im obigen Sinn verleiht außerdem der ansonsten unhaltbaren Annahme eines Kurzzeitrahmens, wie er im Rahmen mancher Design-Modelle vertreten wird, für die Diversifikation der Vogelgruppen Plausibilität.
| |
|
Die Familie der Honigfresser (Meliphagidae)
 |
| Abb. 1: Chaetoptilus angustipluma, eine der aus der Familie der Honigfresser ausgelagerten Arten. (Wikimedia, Public Domain) |
|
Einführung. Die Familie der Honigfresser (Meliphagidae) aus der Ordnung der Sperlingsvögel (Passeriformes) umfasst 182 Arten. Innerhalb dieser Ordnung werden die Honigfresser der Unterordnung der Singvögel (Passeri) zugerechnet. Ihren eingängigen Namen verdanken sie ihrer Ernährung, die sich größtenteils aus Pollen und Nektar zusammensetzt. Fleischer et al. (2008) nennen als Anpassungen an diese Ernährungsweise die lange, einrollbare, gegabelte und mit Fransen ausgestattete Zunge, sowie den langen – häufig abwärts gekrümmten – Schnabel (vgl. Abb. 1 und 2). Sie sind vor allem in den Bäumen Australiens und Neuguineas anzutreffen, aber durch einige Gattungen (etwa Myzomela, Foulehaio und Gymnomyza) auch auf vielen Inseln im südpazifischen Raum beheimatet. Bisher wurden den Honigfressern auch einige Arten auf Hawaii zugeordnet, auf die im Verlauf des Artikels jedoch separat eingegangen wird.
„Verluste“. Nicht wenige Honigfresser-Arten sind in der jüngeren Vergangenheit komplett ausgestorben oder gelten zumindest als stark bedroht. Doch die Meliphagidae hatten noch anderweitig „Verluste“ zu verzeichnen. So wurde beispielsweise die auf Neuseeland endemische Art Notiomystis cincta aufgrund molekularer Untersuchungen ausgegliedert und in eine eigene Familie (Notiomystidae) gestellt (Driskell et al. 2007; Ewen et al. 2006). Allerdings hatte die Morphologie von N. cincta schon früher Anlass für den Verdacht geliefert, die Art gehöre zwar in die Nähe der Honigfresser, jedoch nicht in die Familie selbst (Higgins et al. 2001).
|
|
Die Hawai-Honigfresser
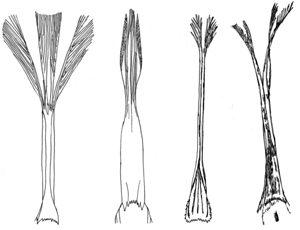 |
| Abb. 2: Gefranste Zungen zweier Meliphagiden (a, b) und zweier Moho-Arten (c, d). Nach den hier vorgestellten molekularen Untersuchungen muss diese spezielle Form der Zunge konvergent entstanden sein. a Meliphaga fasciogularis, b Myzomela sclateri, c Moho nobilis, d Moho braccatus. (Nach Fleischer et al. 2008) |
|
Die Arten. Nun wurde jedoch eine Ausgliederung vorgenommen, welche ein weit größeres Ausmaß und bedeutendere Folgerungen hat und außerdem völlig unerwartet kam. Betroffen sind der Schmalfederhonigfresser (Chaetoptila angustipluma, Abb. 1) und alle vier Arten der Krausschwänze (Moho apicalis [† ca. 1837], M. bishopi [... ca. 1904], M. nobilis [... ca. 1943], M. braccatus [... ca. 1987]). Da C. angustipluma ebenfalls als ausgestorben gilt, müssen taxonomische Untersuchungen auf Museumsmaterial zurückgreifen. Während für das Erklären des Aussterbens von C. angustipluma natürliche Ursachen ausreichend sein dürften, spielte es bei der Gattung Moho auch eine Rolle, dass ihre gelben Federbüschel für die Kleidung der Adelsstände begehrt waren.
Beziehung zu anderen Honigfressern. Die fünf Arten ernährten sich wie die Honigfresser von Nektar und besaßen auch die genannten Anpassungen. Anders als bei N. cincta war ihre Einordnung als Honigfresser nie in Frage gestellt worden (vgl. beispielsweise Merrem 1786; Lesson 1831; Perkins 1903; Amadon 1950). Wie Fleischer et al. (2008) versichern, hätten Taxonomen nie daran gezweifelt, dass Chaetoptila und Moho den Honigfressern zuzuordnen seien. Die einzige Uneinigkeit bezog sich darauf, ob die Hawaii-Honigfresser auf eine einzige Besiedlung zurückzuführen, also monophyletisch, und welche Honigfresser-Gattungen ihre nächsten Verwandten seien.
|
|
Molekulare Untersuchungen
Analysen mitochondrialer DNA (mtDNA) von im 19. Jahrhundert gesammelten Individuen der fünf Arten konnten zwar die Monophylie des aus Moho und Chaetoptila bestehenden Artkomplexes belegen, brachten jedoch auch eine äußerst überraschende Erkenntnis: Die molekularen Daten lieferten keine Hinweise darauf, dass sie überhaupt in die Familie der Honigfresser gehören. Dasselbe Ergebnis wurde auch mit zwei Kern-kodierten Genbereichen für eine Auswahl der fünf Arten erzielt. Darüber hinaus gehören Moho und Chaetoptila nach diesen Resultaten nicht einmal zur selben Radiation wie die Honigfresser, sondern sind der Teilordnung Passerida (Barker et al. 2004) zuzurechnen, sind also mit ihrer bisherigen Familie nur sehr entfernt verwandt. Am nächsten stehen sie nun vielmehr den Seidenschwänzen (Bombycillidae), dem Trauerseidenschnäpper (teilweise als eigene Familie Ptilogonatidae) und dem Palmschwätzer (Dulidae). Die Vögel dieser Familien ernähren sich jedoch nur von Früchten und Insekten und sind dementsprechend auch Moho/Chaetoptila auf der morphologischen Ebene unähnlich. Daher führen Fleischer et al. (2008) für die beiden Gattungen die neue Familie „Mohoidae“ ein. Ein Merkmal, das die Mohoidae mit den genannten nahen Verwandten teilen, das angesichts der großen Übereinstimmungen mit den anderen Honigfressern aber wenig beachtet wurde, ist jedoch, dass sich all diese Arten nomadisch in Schwärmen bewegen. Lovette (2008) meint, dieses Verhaltensmuster hätte die Besiedlung Hawaiis vom Festland aus (und eben nicht durch „island-hopping“ über den Südpazifik durch Honigfresser) ermöglicht.
|
|
Die Konvergenzen
Die gemeinsamen Merkmale, mit welchen die Mohoidae und die Meliphagidae „Generationen von Taxonomen in die Irre führten“ (Fleischer et al. 2008, 1928), stellen also keine homologen Eigenschaften dar und müssen folglich konvergent entstanden sein. Das ist besonders insofern erstaunlich, als die Gemeinsamkeiten zwischen den beiden Familien über die bereits erwähnten Schnäbel und Zungen hinausgeht. Noch viele andere – mit dem Nahrungserwerb zusammenhängende – Merkmale sind betroffen, wie etwa die Gestalt der Füße und Beine, welche für das Erreichen von Blüten von Vorteil sind. Außerdem ist noch der unabhängig evolvierte Schutz der Nasenhöhlen vor Pollen zu nennen. Auch in Verhalten, Federkleid (etwa der gelbe Federbüschel, s.o.; Lovette 2008) und Gesang tauchen deutliche Parallelen auf. Fleischer et al. (2008) berichten außerdem, dass die Ähnlichkeiten sich sogar noch auf morphologische Untergruppen erstrecken: Mindestens drei Formtypen („morphotypes“) der Mohoidae finden sich auch bei den Honigfressern. Fleischer et al. (2008) führen diese weitreichenden Konvergenzen auf ähnliche Selektionsdrücke zurück, die mit der Verteidigung von Nektar-Quellen in Verbindung stehen und machen die bemerkenswerte Feststellung: „Diese Konvergenzen sind derart durchgehend, dass es ohne die molekularen Sequenz-Daten wohl nie möglich gewesen wäre, herauszufinden, dass die nächsten Verwandten der Hawaii-Abstammungslinie die Seidenschwänze und ähnliche sind“ (S. 1927).
|
|
Zeitrahmen und Diversifikation
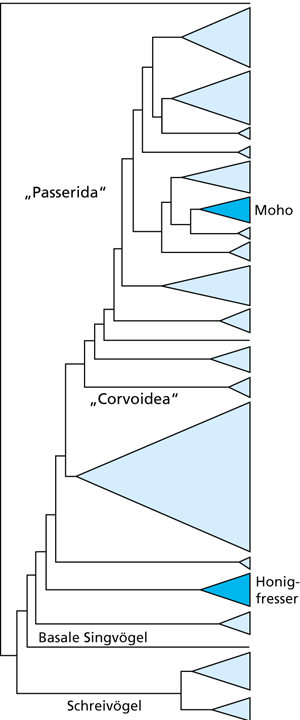 |
| Abb. 3: Schematisches Cladogramm aus 190 Sequenzen von bis zu 1544 Basenpaaren des RAG-1-Gens der Singvogelgruppen. Es zeigt, dass die Gattung Moho genetisch nicht näher mit den Honigfressern verwandt ist (farblich jeweils hervorgehoben). (Stark vereinfacht nach Fleischer et al. 2008) |
|
Die herkömmliche Deutung. Fleischer et al. (2008) betonen einerseits das Ausmaß der Konvergenz, meinen andererseits jedoch auch: „[M]an muss den Zeitrahmen berücksichtigen, der vergangen ist, seit sich die Abstammungslinie von den Vorfahren auf dem Festland getrennt hat.“ Die Autoren schätzen, dass 14 bis 17 Millionen Jahre (bzw. 10-20 nach einem anderen Verfahren) vergangen sind, seit Moho und der nächste Verwandte auf dem Festland, der Trauerseidenschnäpper (Phainopepla nitens aus der Familie der Bombycillidae oder alternativ als eigene Familie Ptilogonatidae), ihren letzten gemeinsamen Vorfahren hatten. Nach Auffassung der Autoren reicht diese Zeit gut aus, damit die hier beschriebenen Anpassungen konvergent evolutiv hätten verwirklicht werden können. Diese Daten würden außerdem belegen, dass es sich bei den Mohoidae um die älteste Vogelgruppe auf Hawaii handelt (Fleischer & McIntosh 2001; Price & Claque 2002) und – falls die weniger weit zurückreichenden Schätzungen korrekt wären – dass ihr Auftauchen mit dem Auftreten von vogelbestäubten Pflanzen zusammenfallen würde (Givnish et al. 2009; Price & Claque 2002).
Alternative Deutung durch eine teleologische Perspektive. Alternativ zur Annahme der konvergenten Annäherung der beiden Vogelfamilien könnte man auch von einem gemeinsamen – in dieselbe Richtung weisenden – genetischen Potential ihrer Vorfahren ausgehen. Wie der Bezug auf eine „Richtung“ bereits deutlich macht, ist ein solches Potential natürlich eng mit teleologischen Implikationen verbunden: Komplexe Voranpassungen an spätere Bedingungen können nur schlecht auf Zufall zurückgeführt werden und legen eine zielsetzende Instanz nahe. Diese könnte eine simulierte Zukunft vorwegnehmen und wäre fähig, dementsprechend zu handeln. Daher kann sie als „Intelligenz“ bzw. als „Designer“ und das Produkt ihres Handelns als „Design“ bezeichnet werden. Dementsprechend bietet sich für den hier vorgelegten Befund ein Interpretationsrahmen an, der nicht nur auf ungerichtete Prozesse wie die natürliche Selektion zurückgreift, sondern teleologische Aspekte beinhaltet. Beispielhaft für eine Deutungsmöglichkeit in diesem übergeordneten Rahmen sei das Konzept des „Frontloadings“ nach Gene (2007) genannt, welches postuliert, bereits die ersten Lebensformen seien so ausgestattet worden, dass ihre zukünftige Evolution in eine bestimmte Richtung gehen würde. Solche Konzepte, die Bezug auf die Identität des Designers nehmen (auf die „W-Fragen“, z.B. die Frage nach dem „Wie“, nach der Methode des Designens, eingehen), stellen mehr oder weniger weit ausgearbeitete Modelle spezifischen Designs (= SD-Modelle) dar, da ihre Erwartungen von der Wahl der spezifischen Eigenschaften des Urhebers abhängen.
Kurzzeitrahmen. SD-Modelle, welche sich einem Kurzzeitrahmen für die Interpretation der naturwissenschaftlichen Befunde zur Geschichte des Lebens verschrieben haben, wie es etwa bei einer biblisch begründeten Schöpfungslehre der Fall ist (Junker 2004; 2005), haben keine 10 Millionen Jahre zur Verfügung, wie sie für die hier diskutierte konvergente Evolution im konventionellen Zeitrahmen veranschlagt werden. Sollen die hier
 |
Die Konvergenzen sind derart
ausgeprägt, dass sie ohne die
molekularen Untersuchungen nicht
entdeckt worden wären. |
 |
|
beschriebenen Merkmalskomplexe jedoch nicht durch ungerichtete Prozesse parallel entstanden sein, erübrigt sich auch die Notwendigkeit langer Zeiträume. Das gemeinsame Design könnte bereits in einer gemeinsamen Stammform angelegt worden sein, oder zwei verschiedenen – stammesgeschichtlich nicht verbundenen – Ausgangspopulationen verliehen worden sein. Kurz gesagt: Tiefe Homologie statt Konvergenz! Homologie entweder im Sinne eines gemeinsamen, aber über weite Teile der Stammesgeschichte latenten Erbes oder mit der Bedeutung eines gemeinsamen zugrundeliegenden Planes. Letzteres entspricht dem Homologieverständnis, wie es ursprünglich vertreten wurde (vgl. Junker & Scherer 2006, 170).
Als Argument gegen eine unabhängige Entstehung durch ungerichtete Prozesse und für ein gemeinsames Design könnte man anmerken: Es scheint zweifelhaft, dass für derart nahrungsunspezifische Merkmale, wie etwa den Gesang und das Federkleid, vergleichbare Selektionsdrücke formulierbar sind.
Ein solches homologes Anpassungspotential sollte, anders als etwa in manchen Beiträgen bei Scherer (1993), nicht nur auf die Existenz verschiedener Allele in der Ausgangspopulation zurückgeführt werden, sondern auch latente Genkomplexe, spezielle genetische Mechanismen zur Steigerung der Rate positiver Mutationen und zielgerichtete Mikroevolution in Betracht ziehen (vgl. Heilig 2008a). Dementsprechend stellt sich die Frage, ob die von den polyvalenten Stammformen ausgehenden Radiationen tatsächlich adaptiver Natur waren und auf der Grundlage bekannter mikroevolutiver Prozesse erfolgten (vgl. Scherer 1993) oder ob die Radiationen nicht-adaptiv waren (Wood 2005, 187-197; Eck [1988] spricht in dieser Hinsicht von „evolutiver Radiation“ im Kontrast zu „adaptiver Radiation“), was sie zeitlich praktisch beliebig schnell hätte ablaufen lassen können. Dabei ist interessant, dass auch aus der nicht-schöpfungswissenschaftlichen Literatur zu den Singvögeln (Gattung Junco) bereits Artbildung berichtet wurde, die in vielleicht nur 1300 Jahren (oder sogar noch weniger Zeit: vgl. Heilig 2008b) erfolgte (Milà et al. 2007).
|
Literatur
- Amadon D (1959)
- The Hawaiian honeycreepers (Aves: Drepaniidae). Bull. Am. Mus. Nat. Hist. 95, 157-262.
- Barker FK, Cibois A, Schikler P, Feinstein J & Cracraft J (2004)
- Phylogeny and diversification of the largest avian radiation. Proc. Natl. Acad. Sci. USA 101, 11040-11045.
- Driskell AC, Christidis L, Gill B, Boles WE, Barker FK & Longmore NW (2007)
- A new endemic family of New Zealand passerine birds: adding heat to a biodiversity hotspot. Australian J. Zool. 55, 1-6.
- Ewen JG, Flux I & Ericson PGP (2006)
- Systematic affinities of two enigmatic New Zealand passerines of high conservation priority, the hihi or stitchbird Notiomystis cincta and the kokako Callaeas cinerea. Mol. Phylog. Evol. 40, 281-284.
- Eck S (1988)
- Gesichtspunkte zur Art-Systematik der Meisen (Paridae) (Aves). Zool. Abh. Staatl. Mus. Tierk. Dresden 43, 101-143.
- Fleischer RC & McIntosh CE (2001)
- Molecular systematics and biogeography of the Hawaiian avifauna. Studies Avian Biol. 22, 51-60.
- Fleischer RC, James HF & Olsen SL (2008)
- Convergent evolution of Hawaiian and Australo-Pacific Honeyeaters from distant songbird ancestors. Curr. Biol. 18, 1927-1931.
- Gene M (2007)
- The Design Matrix. A Consilience of Clues. Arbor Vitae Press.
- Givnish TJ, Millam KC, Mast AR, Paterson TB, Theim TJ, Hipp AL, Henss JM, Smith JF, Wood KR & Sytsma KJ (2009)
- Origin, adaptive radiation and diversification of the Hawaiian lobeliads (Asterales: Campanulaceae). Proc. Biol. Sci. 276, 407-416.
- Heilig C (2008a)
- Ruineneidechsen: Makroevolution oder Polyvalenz? Rapide Anpassung, Makroevolution und Hinweise auf programmierte Variabilität bei Podarcis sicula (Sauria: Lacertidae). Stud. Int. J. 15, 76-88.
- Heilig C (2008b)
- Neues zur explosiven Artbildung. Stud. Int. J. 15, 47-48.
- Higgins PJ, Peter JM & Steele WK (eds., 2001)
- Handbook of Australian, New Zealand and Antarctic Birds Volume 5: Tyrant-Flycatchers to Chats. Melbourne.
- Junker R (2004)
- „Harter Kern“ und Hilfshypothesen von Forschungsprogrammen in der Schöpfungsforschung.
- Junker R (2005)
- Wissenschaft im Rahmen des Schöpfungsparadigmas.
- Junker R & Scherer S (2006)
- Evolution. Ein kritisches Lehrbuch. Gießen.
- Lakatos I (1974)
- Falsifikation und die Methodologie wissenschaftlicher Forschungsprogramme. In: Lakatos I & Musgrave A (Hg) Kritik und Erkenntnisfortschritt. Abhandlungen des Internationalen Kolloquiums über die Philosophie der Wissenschaft, Bd. 4. Braunschweig, S. 89-189.
- Lesson RP (1831)
- Traité d’Ornithologie. Paris.
- Lovette IJ (2008)
- Convergent evolution: Raising a family from the dead. Curr. Biol. 18, R1132-R1134.
- Merrem B (1786)
- Avium rariorum et minus cognitarum, Icones et Descriptiones. Lipsiae.
- Milà B, McCormack JE, Castenada G, Wayne RK & Smith TB (2007)
- Recent postglacial range expansion drives the rapid diversification of a songbird lineage in the genues Junco. Proc. R. Soc. B 274, 2653-2660.
- Perkins RCL (1903)
- Fauna Hawaiiensis: Vertebrata. Cambridge University Press.
- Price JP & Clague DA (2002)
- How old is the Hawaiian biota? Geology and phylogeny suggest recent divergence. Proc. Biol. Sci. 269, 2429-2435.
- Scherer S (Hg., 1993)
- Typen des Lebens. Berlin.
- Wood TC (2005)
- A Creationist Review and Preliminary Analysis of the History, Geology, Climate, and Biology of the Galápagos Islands. Eugene.
|
|  |
