 |
|
| |
 |
| Abb.1: Arbeiterin der stachellosen Biene Proplebeia dominicana als Inkluse in Dominikanischem Bernstein. Sie transportiert Pollinien und ist mutmaßliche Bestäuberin einer als Meliorchis caribea gen. et sp. nov. beschriebenen Orchidee. Abdruck mit freundlicher Genehmigung von S.R. Ramírez. |
|
Orchideen (Orchidaceae) stellen die artenreichste Familie unter den Angiospermen (bedecktsamigen Blütenpflanzen) dar. Sie finden sich mit Ausnahme von arktischen Gebieten überall auf der Erde (Kosmopoliten). Die auffällig verschiedenartig ausgeprägten Blüten sind in oft sehr markanter Weise an unterschiedlichste Bestäuber wie Bienen, Hummeln, Schmetterlinge, Käfer, Vögel, Fledermäuse und Frösche angepasst. Fossile Hinweise auf Orchidaceae sind extrem selten und die wenigen Beispiele häufig umstritten.
Ramírez et al. (2007) haben eine mit Orchideen-Pollinien beladene stachellose Biene (Proplebeia dominicana) beschrieben, die in Dominikanischem Bernstein eingeschlossen ist (Abb. 1). (Ein Pollinium ist ein kleines Tröpfchen aus verklebtem Pollen, das als gesamtes Paket auf den Bestäuber übertragen wird.) Bernstein (fossiles Harz) wird in der Dominikanischen Republik in Sedimenten aus dem Miozän gefunden, für die Iturralde-Vinent & MacPhee (1996) ein Alter von 15-20 Millionen Jahren angegeben haben. Die Autoren sehen in ihrer Arbeit das erste eindeutige Fossil der Orchidaceae sowie einen schönen Beleg für die Wechselwirkung zwischen Pflanze und Bestäuber.
Aufgrund von Vergleichen mit Pollinien aus Herbarien beschreiben Ramírez und Mitarbeiter eine neue Orchideenart: Meliorchis caribea gen. et sp. nov., die sie dem Subtribus Goodyerinae zurechnen. Die Position der Pollinien auf dem Rücken (Mesoscutellum) der Biene erlaubt den Autoren Rückschlüsse auf den Blütenaufbau. Heute wachsende Vertreter der Goodyerinae übertragen die Pollinien nämlich auf die Mundwerkzeuge der bestäubenden Bienen. Also muss die fossile Biene im Gegensatz zu ihren heutigen Verwandten bei Meliorchis caribea mit dem gesamten Insektenkörper in die Blüte eingedrungen sein.
Um die Verwandtschaft von Meliorchis caribea aufzuklären, berechneten die Autoren anhand von 25 morphologischen Merkmalen Stammbäume. Die eingesetzten Programme ergaben 129 gleichwertige Stammbäume, wobei die fossile Orchidee immer unter die Goodyeridae eingruppiert wurde. In einer weiteren Berechnung versuchten die Autoren zu ermitteln, wann die Orchidee gewachsen ist, von der die heutigen Orchideen abstammen. Dazu verwendeten sie DNA-Sequenzen aus Plastiden von 55 Orchideen und nutzten die fossilen Befunde, um eine molekulare Uhr zu kalibrieren. Die Rechnung ergab, dass die „Ur-Orchidee“ in der späten Kreide gewachsen ist, vor 76 bis 84 Millionen Jahren. Damit – so die Autoren – sei die Hypothese einer jungen Entstehung (Eozän oder jünger) der Orchideen zugunsten einer älteren (späte Kreide) abgewiesen und die Verbreitung der Orchideen mit Schwerpunkt in den tropischen Klimazonen durch den Austausch zwischen den tropischen Kontinenten in der späten Kreide plausibel erklärt.
Einmal mehr hat sich Bernstein als herausragendes Fenster in die Erdgeschichte erwiesen und einen faszinierenden Einblick in ökologische Zusammenhänge zwischen Orchideen und den sie (vermutlich) bestäubenden Bienen erlaubt. Ob die von Ramírez et al. (2007) angestellten Überlegungen und Berechnungen zum Ursprung der Orchideen überzeugen, hängt nicht zuletzt von der Akzeptanz der hinter den verwendeten Algorithmen stehenden Modelle und deren Randbedingungen ab. Die Hypothese eines jungen Ursprungs der Orchideen und die Vorstellung, dass deren ausgeprägte Vielfalt und Variabilität eine Pflanzenfamilie in der Phase der Etablierung zeigt (Schmid & Schmid 1977), sind mit diesem Befund in Frage gestellt.
[Iturralde-Vinent ME & MacPhee RDE (1996) Age and paleogeography of Dominician amber. Science 273, 1850-1852; Ramírez SR, Gravendeel B, Singer RB, Marshall CR & Pierce NE (2007) Dating the origin of the Orchidaceae from a fossil orchid with its pollinator. Nature 448, 1042-1045; Schmid R & Schmid MJ (1977) Fossil history of the Orchidaceae. In: Arditti J (ed.) (1977) Orchid Biology: Reviews and Perspectives Vol. 1. London, pp 17-45.]
|

 |
| Abb.1: Weichkäfer (Cantharidae) in fossilem Harz aus Birma. In Verteidigungssituation (siehe fremdes Fühlerfragment am Hinterleib des Käfers) sind an beiden Seiten ausgesonderte Sekrettröpfchen erkennbar. Abdruck mit freundlicher Genehmigung von G.O. Poinar Jr. |
|
In seiner empfehlenswerten, mit ausgezeichneten Bildern illustrierten Monographie „In love for Insects“ hat Thomas Eisner (2003) seine umfangreichen Erfahrungen aus Beobachtungen und Untersuchungen von Insekten zusammengetragen. Vielen von ihnen hat der enthusiastische Entomologe gemeinsam mit seinem Freund und Chemiker J. Meinwald chemische Geheimnisse entlockt, mit denen sie beispielsweise ihre Fortpflanzungspartner anlocken oder sich vor Fressfeinden schützen.
Einen Weichkäfer (Familie Cantharidae, Abb. 1), der sich vermutlichen in Erwartung eines Angriffs mit chemischen Waffen zur Wehr setzt, beschreiben Poinar et al. (2007). Die Szenerie wird aber nicht irgendwo auf der Wiese beobachtet und dokumentiert, sondern sie ist in fossilem Harz aus Birma festgehalten und konserviert. Vom vermuteten Angreifer ist in dem erhaltenen Stück fossilen Harzes nur ein Teil eines Fühlers zu erkennen. Die Autoren spekulieren, dass es eine Schabe gewesen sein könnte; diese Allesfresser sind in Bernstein aus Birma mehrfach beschrieben.
Das fossile Harz wurde in einem Braunkohleflöz gesammelt, das in Sandstein-Kalk Sedimenten eingelagert ist. Aufgrund paläontologischer Befunde wird die Lagerstätte dem oberen Alb der frühen Kreide zugeordnet (Cruichshank & Ko 2003), was einem radiometrischen Alter von rund 100 Millionen Jahren entspricht. Poinar et al. (2007b) haben Hinweise auf Auraucariaceae als Harzlieferpflanzen für den Bernstein aus Birma veröffentlicht. Die bisher ältesten fossilen Hinweise auf Arten der Cantharidae stammen aus baltischem Bernstein (Oligozän/Eozän, ca. 30-40 Millionen Jahre; Weitschat & Wichard 1998).
Der Käfer mit einer Länge von ca. 4,5 mm ist in einem klaren Stück fossilen Harzes (7 x 4 x 2 mm) eingeschlossen und gut erhalten. Aus den ersten Segmenten des Abdomen (Hinterleib) sind auf beiden Seiten Bläschen erkennbar, die von dahinter liegenden Drüsen ausgepresst werden. Die massivste Sekretion (Länge: 1,4 mm) erfolgt an der Stelle, an der der Käfer in Kontakt mit dem fremden Fühler ist. An der Kontaktstelle haftet Sekret am Fühler und ein großer Sekretstrang ist über dem Thorax verteilt.
Poinar et al. (2007) erwähnen anhand von Literaturzitaten, dass bei Käfern (Coleoptera) chemische Abwehrmechanismen mindestens 30 mal entwickelt wurden – ein beeindruckendes Beispiel von Konvergenz. Bei mikroskopischer Betrachtung der Sekrete kann man Kristalle erkennen, deren chemische Analyse jedoch nur auf Kosten der Zerstörung des Fossils möglich wäre. In mehreren Arbeiten haben Eisner und Meinwald – sowie auch weitere Arbeitsgruppen (zitiert in Poinar et al. 2007) – gezeigt, dass Weichkäfer der Gattung Chauliognathus verschiedene Carbonsäuren, Triglyceride und Glycerinester durch Drüsen ausscheiden können.
Heute scheinen chemische Verteidigungsstrategien bei Insekten weit verbreitet zu sein (Eisner 2003). Das von Poinar et al. (2007) beschriebene Fossil belegt, dass diese Option bereits in der unteren Kreide von Weichkäfern eingesetzt wurde. Angesichts der geringen chemischen Stabilität fossiler Harze kann man allerdings die Frage stellen, warum Bernstein aus der Kreide, wenn diese einen Zeitraum von vor über 100 Millionen Jahren beschreibt, überhaupt noch da ist. Aufgrund chemischer Erfahrungen ist dies zumindest erstaunlich.
[Cruichshank RD & Ko K (2003) Geology of an amber locality in the Hukawng Valley, northern Myanmar. J. Asian Earth Sci. 21, 441–455; Eisner T (2003) For Lofe of Insects. Cambridge, Mass.; Poinar GO, Marshall CJ & Buckley R (2007a) One hundred million years of chemical warfare by insects. J. Chem. Ecol. 33,1663-1669; Poinar GO, Lambert J B & Wu Y (2007b) Araucarian source of fossiliferous Burmese amber: spectroscopic and anatomical evidence. J. Bot. Res. Inst. Texas 1, 449–455; Weitschat W & Wichard W Atlas der Pflanzen und Tiere im Baltischen Bernstein. München 1998; S. 162-167.]
[Cruichshank RD & Ko K (2003) Geology of an amber locality in the Hukawng Valley, northern Myanmar. J. Asian Earth Sci. 21, 441–455; Eisner T (2003) For Lofe of Insects. Cambridge, Mass.; Poinar GO, Marshall CJ & Buckley R (2007a) One hundred million years of chemical warfare by insects. J. Chem. Ecol. 33,1663-1669; Poinar GO, Lambert J B & Wu Y (2007b) Araucarian source of fossiliferous Burmese amber: spectroscopic and anatomical evidence. J. Bot. Res. Inst. Texas 1, 449–455; Weitschat W & Wichard W Atlas der Pflanzen und Tiere im Baltischen Bernstein. München 1998; S. 162-167.] |
|
Vor rund 10 000 Jahren soll sich die letzte Eiszeit ihrem Ende zugeneigt haben. Im Zuge der damit einhergehenden Wiederbesiedlung der freiwerdenden Landflächen soll auch der Singvogel Junko (Gattung Junco) eine bemerkenswerte Diversifikation erfahren haben, wie Milà et al. (2007) durch eine neue Studie eindrucksvoll belegen. Interessanterweise benötigte der Vogel dafür „nicht einmal eine isolierte Nische, sondern nutzte gleich einen ganzen Kontinent“, wie in Spektrumdirekt (2007) berichtet wird. Dies widerlegt die Allgemeingültigkeit der bisher vertretenen Annahme bezüglich der nordamerikanischen Vogelvielfalt, „dass die Diversifikation erfolgte, als die Populationen über lange Zeiträume in Refugien isoliert waren, die wahrscheinlich einen bis mehrere volle Eiszeitzyklen umfassten“ (Milà et al. [2007]; vgl. Johnson & Cicero 2004; Weir & Schluter 2004). Vielmehr rückt nun auch das sich an das Pleistozän anschließende Holozän ins Blickfeld des Interesses.
 |
| Abb.1: Winterammer Junco hyemalis (GNU Freie Dokumentationslizenz) |
|
Vom südlichen Mesoamerika ausgehend soll der Junko (bzw. die Junko-Art Junco phaenotus, „Rotrückenjunko“, „yellow-eyed junco“) die freiwerdenden Gebiete besiedelt und dabei seine heutige Vielfalt erreicht haben. Besonders hervorzuheben ist die dabei neu entstandene Art der Winterammer (Junco hyemalis; „dark-eyed junco“; Abb. 1), welche weiter nördlich wiederum eine beeindruckende Vielfalt erreichte und mittlerweile in fünf verschiedenen Unterarten auftritt, die lange Zeit sogar für eigene Arten gehalten wurden (Spektrumdirekt 2007). Berichte von stabilen Hybridisierungszonen stellten diese Arttrennung jedoch in Frage (Nolan et al. 2002). Dies gilt jedoch nicht für alle fünf Formen: Für zwei Unterarten (J. oreganus und J. caniceps) gibt es bereits Hinweise auf ein Fehlen von Kreuzungen in gemeinsam besiedelten Gebieten, was als Indiz für bereits stattgefundene sympatrische (im gleichem Gebiet erfolgte) Artbildung gilt (Johnson & Cicero 2004).
Die Ergebnisse sind auch insofern interessant, als sie erneut belegen, dass das Erreichen einer großen Vielfalt in kurzer Zeit durchaus möglich ist, wenn eine Stammart mit entsprechend großem genetischen Potential vorliegt und diese einen neuen, frei werdenden Lebensraum besiedelt (siehe auch Stephan 2002).
Wie viel Zeit ist für die Artbildung zu veranschlagen? Laut molekularer Uhr und genetischen Daten aus mitochondrialen Kontrollregionen ergibt sich für die Entstehungsdauer der neuen Junko-Art eine Zeitspanne von 1300-3800 Jahren, was die Autoren als „außergewöhnlich“ bezeichnen. Andere Untersuchungen bestätigten diese Zahl als Untergrenze und lassen eine Addition von bis zu 7000 Jahren offen. Dem unteren Wert von 1300 Jahren liegt eine Mutationsrate von 15% pro Million Jahre zugrunde. Aus der Klasse der Vögel sind jedoch auch wesentlich höhere Mutationsraten bekannt: Beispielsweise 21% bei Gänsen (Quinn [1992]) und gar 30% bei Pinguinen (Lambert et al. 2002). Eine solche Mutationsrate hätte zu noch größeren Diversifikationsraten geführt. In diesem Zusammenhang ist des weiteren zu bedenken, dass die Dauer des Holozän, in dem sich die Disversifikation des Rotrückenjunkos komplett vollzogen haben soll, laut Menting (2002) um 50% und mehr gekürzt werden muss, um die Daten realistisch verstehen zu können.
Die durchschnittliche Zeitspanne für Artbildungen in der Klasse der Vögel wird auf 0,1-0,2 Millionen Jahren geschätzt. Dies liegt unter anderem darin begründet, dass das Pleistozän weiterhin eine wichtige Rolle für die Vogeldiversifikation spielen soll, auch wenn die Ergebnisse der hier vorgestellten Studie den Fokus mehr in Richtung Holozän verschiebt. Für das Pleistozän werden im konventionellen Zeitrahmen 1,8 Millionen Jahre veranschlagt. Sollte sich die von Brandt (2006) vorgeschlagene drastische Verkürzung auf nur mehrere Tausend Jahre weiter belegen lassen, so müsste auch der Zeitrahmen für die Artaufspaltungen bei den Vögeln entsprechend deutliche Kürzungen erfahren.
[Johnson NK & Cicero C (2004) New mitochondrial DNA data affirm the importance of Pleistocene speciation in North American birds. Evolution 58, 1122-1130; Lambert DM, Ritshie PA, Millar CD, Holland B, Drummond AJ & Baronoi C (2002) Rates of evolution inancient DNA from Adélie penguins. Science 295, 2270-2273; Menting G (2002) Die kurze Geschichte des Waldes. Plädoyer für eine drastische Kürzung der nacheiszeitlichen Waldgeschichte. Andernach; Milà B, McCormack JE, Castenada G, Wayne RK & Smith TB (2007) Recent postglacial range expansion drives the rapid diversification of a songbird lineage in the genus Junco. Proc. R. Soc. B 274, 2653-2660; Nolan VJ, Ketterson ED, Cristol DA, Rogers CM, Clotfelter ED, Titus RC, Schoech SJ & Snajdr E (2002) Dark-eyed junco (Junco hyemalis). In: Poole A & Gill F (2002) The birds of North America. Vol. 716, pp. 1-44; Quinn TW (1992) The genetic legacy of mother goose. Phylogeographic patterns of lesser snow goose Chen caerulescens caerulescens maternal lineages. Mol. Ecol. 1, 105-117; Spektrumdirekt (2007) Artbildung im Zeitraffer. Spektrum direkt – die Wissenschaftszeitung im Internet, 30. August; Stephan M (2002) Der Mensch und die geologische Zeittafel. Warum kommen Menschenfossilien nur in den obersten geologischen Schichten vor? Holzgerlingen; Weir JT & Schluter D (2004) Ice sheets promote speciation in boreal birds. Proc. R. Soc. B 271, 1881-1887.]
|
|
Diese vielleicht etwas trivial erscheinende Einsicht hat Kobayashi & Koshima (1997) immerhin einen kleinen Artikel in Nature ermöglicht. Die Forscher verglichen menschliche Augen mit denen von 94 anderen Primaten. Die sogenannte Lederhaut, also der Bereich, der die Iris umgibt, ist bei Menschen besonders groß. Zudem sind die Augenöffnungen horizontal besonders breit, sie erlauben dadurch größere Augenbewegungen. Menschen sind außerdem die einzigen Primaten mit weißer Lederhaut. Alle diese Faktoren führen dazu, dass man die Blickrichtung beim menschlichen Auge besonders gut erkennen kann. Die Analyse der Blickrichtung ist ein wichtiger Bestandteil der Kommunikation. Die Einfachheit der Blickrichtungsanalyse beim Menschen zeige, wie wichtig diese Art der Kommunikation für Menschen sei und gleichzeitig ist es ein trennendes Merkmal von Menschen zu anderen Primaten.
[Kobayashi H & Kohshima S (1997) Nature 387, 767-768]
|
|
Die für uns eigenartig anmutende Möglichkeit der unabhängigen Augenbewegung bei Chamäleons hat sicher schon jeder einmal im Zoo bewundert. Diese unabhängige Augenbewegung ist aber nicht die einzige Besonderheit. Die stärkste Lichtbrechung wird durch die Hornhaut und nicht durch die Linse bewirkt. Die Linse selbst arbeitet als Zerstreuungslinse. Dadurch gleicht der Augenaufbau mehr einem Teleobjektiv. Wenn man einem Chamäleon beim Sehen zuschaut, so fällt auf, dass immer ein Auge kurz ruhig gehalten und mit dem anderen die Umgebung mit ruckartigen Bewegungen „abgescannt“ wird. Die unabhängige Augenbewegung verbirgt das Chamäleon vermutlich vor Fressfeinden, die normalerweise zwei (sich gleichartig bewegende) Augen erwarten. Die Entfernung zu einem möglichen Beutetier, meist Insekten, wird nicht oder nur selten über Stereosehen gemessen, wie das bei uns der Fall ist, sondern daraus, wie stark das Auge fokussiert ist. Ganz ähnlich wie bei einem Fotoapparat, an dem man ein Objektiv „scharf“ stellt und dann die Entfernung zum Objekt ablesen kann. Ein weiterer Effekt zur Entfernungsmessung kommt hinzu: Durch die besondere Konstruktion hat das Auge einen hohen Parallaxenfehler, d.h. beim Drehen des Auges in der Horizontalen verschieben sich scheinbar die Gegenstände zueinander. Diese scheinbare Verschiebung kann ebenfalls zur Entfernungsmessung benutzt werden. Ist die Konstruktion des Chamäleon-Auges daher einzigartig? Weit gefehlt, ein Fisch namens Sandaal verfügt über genau dieselben Eigenschaften: unabhängige, abwechselnde, ruckartige Augenbewegungen, Teleskop-Effekt und entsprechende Entfernungsmessung. Gefangen werden natürlich keine Insekten, sondern kleine Krebse, die sich in die Nähe des eingegrabenen Fisches (Name!) verirren. Der Forscher Srinivasan (1999) schreibt über diesen verblüffenden Befund in einem Fachartikel mit interessantem Vokabular: „Seltsamerweise produziert [die Evolution] gelegentlich Tiere aus völlig verschiedenen Familien welche sich anscheinend einem gleichartigen Konstruktionsplan annähern. [...] Obwohl sie zu völlig verschiedenen Familien gehören, das Chamäleon ist ein Reptil und der Sandaal ein Fisch, scheinen sie sich einem gemeinsamen Satz von Design-Prinzipien ihrer visuellen Systeme anzunähern.“ Die verblüffende Übereinstimmung führt Srinivasan aber trotz dieses Vokabulars auf umweltbedingte Entwicklungszwänge zurück (vgl. den Artikel über „Design“ von T. Haller & C. Heilig in dieser Ausgabe).
[Srinivasan MV (1999) Nature 399, 305-306]
|
|
Augen sollen im Laufe der Evolution 40mal unabhängig voneinander entstanden sein. Diese Überlegung wurde angezweifelt, weil es sich herausstellte, dass der Hauptschalter zur Augenentwicklung bei Fruchtfliegen und Mäusen der gleiche ist (pax6). Somit wurde ein gemeinsamer Entstehungsursprung der Augen vermutet. Seit dieser Zeit mehren sich aber Beobachtungen neuer Details zur Augenentwicklung bei den verschiedensten Tierarten, die mit einem gemeinsamen Ursprung nicht verträglich sind. Als ein Beispiel seien hier die Augen eines Parasiten mit dem Namen Neoheterocotyle angeführt. Dieses Tier aus der Gruppe der Plattwürmer parasitiert auf Fischen. Die bewimperte Larve hat zwei Paar Augen, am Vorder- und Hinterende. Die Augen am Vorderende bestehen aus zwei, die am Hinterende aus je einer Sinneszelle (Rhabdomer). Interessanterweise besitzt jede Sinneszelle ihre eigene Linse. Diese Linse geht offensichtlich aus einem Mitochondrium der Augenzelle hervor, denn es lassen sich im Elektronenmikroskop Strukturen erkennen, die für diese Zellbestandteile typisch sind. Mitochondrien sind normalerweise die Kraftwerke einer Zelle. Damit bestehen die Lichtsinnesorgane von Neoheterocotyle aus kleinen „Ein-Zell-Augen“, komplett mit Lichtabschirmung von hinten (Pigmentbecher), besagter Mitochondrien-Linse und lichtempfindlichen Teilen, die mit der Funktion einer Retina vergleichbar sind. Es fragt sich, wozu ein so scheinbar einfach gebautes Tier so komplexe Augen benötigt.
[Fernald RD (2006) Science 313, 1914-1918; Rohde K, Watson NA & Chisholm LA (1999) Int. J. Parasitol. 29, 511-519]
|
|
Der genetische Unterschied zwischen Mensch und Schimpanse erweist sich durch weitere Untersuchungen entgegen ursprünglichen Annahmen als zunehmend größer. Die häufig zitierte Differenz zwischen den beiden Primatengenomen von ca. 1% wird mehr und mehr durch neue Befunde in Frage gestellt (vgl. Binder 2007). In einer Analyse der Genome von Rhesusaffe (Macaca mulatta), Hund (Canis familiaris), Ratte (Rattus norvegicus), Maus (Mus musculus), Schimpanse (Pan troglodytes) und Mensch (Homo sapiens) untersuchten Hahn et al. (2007) Datensätze mit 119.746 Genen aus 9.900 Genfamilien aus allen 6 Arten. Alle Genome sind nach dem sogenannten „Schrotschuss-Verfahren“ (nach C. Venter) mindestens 6-fach sequenziert und mindestens zu 96 % untersucht worden. Hahn und Mitarbeiter zeigen, dass (unter evolutionstheoretischen Voraussetzungen) die Geschwindigkeit von Genverlusten und -gewinnen in der Linie der Primaten zunimmt, besonders bei den Großaffen. In verschiedenen Genfamilien ist die Änderung der Kopienzahl von Genen auffällig groß, die Autoren sehen darin den Einfluss natürlicher Selektion. Die Autoren folgern aus den Untersuchungen, dass im Genom des Menschen (seit seiner vermuteten Abspaltung von der Schimpansenlinie vor 5-6 Millionen Jahren) 678 Gene dazugekommen sind, während im Schimpansengenom 740 Gene verloren wurden. Dies bedeutet, dass sich für 6,4% (1.418 von 22.000) aller menschlichen Gene keine direkt vergleichbaren (orthologen) Gegenstücke im Schimpansengenom finden lassen. Es bleibt spannend zu verfolgen, ob sich dieser Trend im genetischen Unterschied mit wachsenden Kenntnissen fortsetzt. Viel bedeutsamer ist jedoch, die Zusammenhänge und Bedeutungen der im Genom niedergelegten Information zu verstehen. Die großen Unterschiede implizieren zudem ernsthafte Fragen nach den Mechanismen der Veränderungen und auch nach deren Triebfeder.
[Binder H (2007) Über den genetischen Unterschied zwischen Mensch und Schimpanse – der „1 %-Mythos“. Stud. Int. J. 14, 77-78; Hahn MW, Demuth JP & Han S-G (2007) Accelerated rate of gene gain and loss in primates. Genetics (published ahead of print, published on 18. October, 2007 as 10.1534/genetics.107.080077); Dolgin E (2007) Evolutionary sprint made us human. http://sciencenow.sciencemag.org/cgi/content/full/2007/1023/2;23.10.2007]
|
|
Der Kleine Beutenkäfer (Aethina tumida, Abb. 1) ist ein Honigparasit, der natürlicherweise in Afrika südlich der Sahara lebt. Für die afrikanischen Unterarten der Honigbiene (Apis mellifera) stellt der Käfer meist keine Gefahr dar. Sowohl die Käfer als auch dessen Eier und Larven werden von den Bienen erkannt und aus dem Stock getragen. Zum Teil werden die Käfer auch in Waben eingesperrt; Wächterbienen sorgen dafür, dass sie nicht entkommen, und andere „mauern“ den Eingang zu. Der Käfer kann manchmal dennoch entkommen, indem er eine Biene beträllert und so einen Fütterungsreflex auslöst. Bei starkem Befall verlassen die Bienen auch ihr Nest. Allerdings wurde der Käfer nun nach Amerika verschleppt und trifft dort auf Bienen europäischer Abstammung. Hier sorgt er für massive Schäden an den Bienenvölkern, bis hin zum Absterben. Untersuchungen zeigten, dass die Abwehrmaßnahmen der europäischen Bienen zwar dieselben sind wie die der afrikanischen, diese aber nicht mit derselben Intensität durchgeführt werden. Je schwächer eine Kolonie ist, um so leichter fällt es dem Käfer, die Abwehrmechanismen zu umgehen. Die Käferlarven leben dann von den Vorräten an Honig und Pollen sowie von der Bienenbrut. Ausgeschlüpfte Käfer stehen dann aber vor dem Problem, dass das Bienenvolk entweder ausgeflogen oder abgestorben ist, und sie müssen sich eine neue Kolonie suchen.
Bienen kommunizieren über Duftstoffe (Pheromone). Der Käfer kann das Alarmpheromon der Bienen riechen und wird davon angelockt. Interessanterweise riecht der Käfer viel niedrigere Konzentration des Alarmstoffes als die Bienen selbst. Schon länger bekannt ist die Tatsache, dass die Käfergruppe, zu denen auch der Kleine Beutenkäfer gehört, Pilze und Hefen übertragen. Neuere Untersuchungen bestätigten das auch für den Kleinen Beutenkäfer, er überträgt die Hefe Kodemaea ohmeri. Honig- und Pollenvorräte, die man mit dieser Hefe infiziert hatte, erzeugten eine höchste Attraktivität nach ca. 7 Tagen. Es zeigte sich, dass die Hefe beginnt, größere Mengen an Alarmpheromon zu erzeugen. Ist es also ersten Beutekäfern gelungen, ein Bienenvolk zu befallen, so wird die Attraktivität dieses Bienenvolkes für weitere Käfer erhöht. Die Bienen werden in der Folge von den Käfern „überrannt“.
Dies ist ein weiteres Beispiel dafür, dass auch Insekten Mikroorganismen zu ihrem Nutzen einsetzen. Ein anderer, vor kurzem bekannt gewordener Fall war bei den Blattschneiderameisen entdeckt worden. Diese „züchten“ in Poren auf ihrer Außenhaut Bakterien, die ein Antibiotikum erzeugen. Dieses Antibiotikum tötet spezifisch einen parasitischen Pilz ab, der auf den Futterpilz-Kolonien der Blattschneiderameisen lebt. Unklar bleibt freilich, durch welche Selektionsdrücke während der Stammesgeschichte solche Beziehungen überhaupt entstanden sein könnten.
[Winkler N (2006) Ameisen – Neue Überraschungen (Teil 1). Stud. Int. J. 13, 38-39; Pirk C (2007) Parasiten – Süßer Käfer. labor&more 3, 54-55; Torto B, Boucias DG, Arbogast RT, Tumlinson JH, Teal PEA (2007) Multitrophic interaction facilitates parasite-host relationship between an invasive beetle and the honey bee. Proc. Natl. Acad. Sci. USA 104, 8374-8378.]
|
|
Dass eine Fliege gezielt eine Futterquelle oder einen Partner anfliegt, ist nicht weiter verwunderlich. Was aber, wenn man eine Fliege ohne äußere Einflüsse in einer dunklen Kammer ohne weitere Reize fliegen lässt? Eigentlich würde man erwarten, dass sie zufällig „herumeiert“. Ein solches Experiment wurde durchgeführt und die Flugbahn der Fliegen aufgezeichnet und per Computer ausgewertet. Zum Erstaunen der Wissenschaftler stellte sich heraus, dass die Flugbahnen nicht zufällig waren. Die Flugbahnen wiesen nichtlineare Komponenten auf, d.h. der Flug wurde von der Fliege in unvorhersagbarer Weise bestimmt, obwohl die Fliegen keinen äußeren Reizen ausgesetzt waren. Das ist insofern erstaunlich, als der gegenwärtige neurowissenschaftliche Konsens postulierte, dass die Quelle für Variabilitäten in Verhaltensmustern das „Rauschen“ sei, welches letztlich dafür sorgt, dass die Variabilität (hier: Flugbahnen) zufällig oder eben stochastisch ist. Bei der Fliege sorgte eine bislang nicht genau identifizierte (innere) Quelle für nichtlineare Abweichungen von rein zufälligen Flugbahnen, welche dadurch völlig unvorhersagbar (chaotisch) wurden. Das heißt, selbst wenn man Tausende von Fliegen beobachtete, könnte man noch nicht einmal im Mittel wahrscheinliche Flugbahnen voraussagen. Warum ist diese Unterscheidung so interessant? Im Falle stochastischer Flugbahnen läge die Quelle im „Außen“, das heißt, dass irgendwelche äußeren Einflüsse als Auslöser in Frage kämen und Verhalten bestimmten. Nichtlineare Flugbahnen zeigen aber, dass es eben nicht der (äußere) Zufall ist, der die Richtung bestimmt, sondern die Flugbahnen von einem „Innen“ bestimmt werden. Das heißt also, dass Entscheidungen selbst einfacher Organismen nicht dem Rauschen (Zufall) unterliegen, sondern kontrolliert werden (wenn auch nichtlinear). Anders ausgedrückt: Gehirne werfen manchmal den Würfel, doch ist dieses Würfeln ein kontrollierter innerer Prozess.
[Maye A, Hsieh C-H, Sugihara G & Brembs B (2007) Order in Spontaneous Behavior. PLoS ONE 2, e443.]
|
|
Pilze treten typischerweise als Fäulnisbewohner (Saprophyten) oder als Parasiten auf. Darüber hinaus haben sich manche Pilze auch auf räuberische Strategien verlegt, um ihre Nahrung zu gewinnen. Fünf unterschiedliche Fangmethoden von fleischfressenden Pilzen sind bisher beschrieben. Manche Pilze produzieren ringförmige Zellen, mit denen sie im Boden lebende Fadenwürmer (Nematoden) und andere Kleinlebewesen fangen, andere versuchen es mit klebrigen Auswüchsen.
Yang et al. (2007) haben jüngst anhand von molekularbiologischen Untersuchungen versucht, die verschiedenen Entwicklungsstufen dieser Methoden aufzuzeigen. Die Autoren sehen in der Entwicklung von Fangtechniken einen gelungenen Beweis für adaptive Evolution – wobei die Entstehung neuer Technik mit dem Begriff „Adaption“ (Anpassung) kaum angemessen beschrieben wird. Abschließend verweisen die Autoren auf einen fossilen Befund von Jannson & Poinar (1968), die Pilzhyphen (Pilzfäden) in Nematoden, als Inklusen (Einschlüsse) in mexikanischem Bernstein (Miozän/ Ologozän; 22,5-26 Millionen Jahre nach radiometrischen Datierungen) beschreiben. Yang et al. 2007 sehen darin einen fossilen Hinweis auf die Zeit, als Pilze ihre Fangmethoden entwickelten, dieser passt allerdings zeitlich nicht mit phylogenetischen Untersuchungen, die auf molekularbiologischen Daten basieren, zusammen.
Schmidt et al. (2007) stellten jüngst Pilzhyphen in fossilem Harz in direkter Verbindung mit Nematoden vor. Der Bernstein stammt aus fossilreichen Ablagerungen in Archingeay/Les-Nouvillers, Südwest-Frankreich, die aufgrund palynologischer Befunde (Pollenanalysen) dem späten Alb (obere Unterkreide; ca. 100 Millionen Jahre) zugeordnet werden.
Das eingeschlossene Pilzgeflecht (Mycel) besteht aus irregulär verzweigten Fäden (Hyphen). Die Ringe bestehen aus einer einzigen Zelle, im Gegensatz zu Fallen-Stukturen heutiger Pilze, die aus drei Zellen bestehen. Die fossilen Pilzringe weisen einen Innendurchmesser von 8 bis 10 µm auf und können sich offenbar leicht von den Hyphen lösen; im fossilen Harz finden sich auch isolierte Ringe. Diese Ringfallen scheinen klebrig gewesen zu sein, zumindest deuten viele an den Ringen haftende Partikel darauf hin. Vermutlich werden Nematoden, die in diesen Ringen gefangen werden, durch Befall mit Hyphen verdaut. In dem Bernsteinstück finden sich mehrere Nematoden in der Nähe von Pilzringen und sie scheinen aufgrund ihres Außendurchmessers die bevorzugte Beute des Pilzes gewesen zu sein. Damit ist also belegt, dass bereits in der frühen Kreide Bodenpilze mit komplexen, hocheffizienten Fangsystemen vorhanden waren. Über 200 Pilzarten füllen heute diese ökologische Nische aus. Der fossile räuberische Pilz kann aufgrund seiner morphologischen Merkmale keinem heute vorkommenden Bodenpilz zugeordnet werden. Fleischfressende Pilze gibt es offensichtlich schon lange als Bodenbewohner und die ökologischen Nischen scheinen im Mesozoikum von anderen, heute nicht bekannten Vertretern besetzt gewesen zu sein.
In Supporting Online Material berichten Schmidt et al. (2007), dass in dem fossilen Harzstück außer dem fleischfressenden Pilzmycel noch 79 Gliederfüßer (zumeist bodenbewohnende Arten) und zahlreiche weitere Mikroorganismen, wie Bakterien und Algen als Einschlüsse enthalten sind. Das ursprüngliche Fundstück (40 x 30 x 20 mm) wurde für die Untersuchung in 31 Teile zerlegt, die jeweils gesondert poliert und mikroskopisch untersucht wurden.
Wieder einmal erweisen sich Einschlüsse in fossilem Harz als hochinformative Quelle für Ökosysteme aus vergangener Zeit. In diesem Falle ist belegt, dass hochkomplexe und spezialisierte Lebensweisen von Pilzen sehr früh – und bisher ohne dokumentierte Vorläuferstrukturen auftauchen.
[Jannson H-B & Poinar Jr GO (1968) Some possible nematophagous fungi. Trans. British Mycol. Soc. 87, 471-474; Schmidt AR, Dörfelt H & Perrichot V (2007) Carnivorous fungi from Cretaceous amber. Science 318, 1743; Schmidt AR, Dörfelt H & Perrichot V (2007) Supporting Online Material: http://www.sciencemag.org/cgi/content/full/318/5857/1743/DC1; Yang Y, Yang E, An Z & Liu X (2007) Evolution of nematode-trapping cells of predatory fungi of the Orbiliaceae based on evidence from rRNA-encoding DNA and multiprotein sequences. Proc. Natl. Acad. Sci. USA 104, 8379-8384.]
|
|
Der erstaunlich geringe Anteil codierender DNA-Sequenzen am Genom (Erbgut) eines Organismus war für manchen Wissenschaftler in der Vergangenheit Anlass, von „Abfall“ („junk“) zu reden – Anteile des Erbguts, die im Laufe der Evolution angesammelt wurden und überflüssig geworden seien. Diese Deutung war zwar nicht unumstritten, aber doch – vor allem in populären Medien – weit verbreitet. Die Anzeichen dafür mehren sich jedoch, dass immer weitere Teile des Genoms von Bedeutung sind. Aber wie kann man analytisch DNA-Abschnitte, die nicht in Proteine übersetzt werden (Translation), überhaupt erkennen? McCallion et al. (2007) haben eine Methode entwickelt, diese im Bereich eines Gens angewendet und dabei überraschende Erfahrungen gemacht.
Die Autoren untersuchten beim Zebrafisch (Danio rerio) die Umgebung des Gens phox2b. Dieses Gen spielt u. a. bei der Entwicklung von Nervenzellen eine Rolle, bei der Stressverarbeitung und im Verdauungssystem. DNA-Fragmente (48 Amplikons) aus diesem Bereich transferierten die Forscher gekoppelt mit dem Gen eines grün fluoreszierenden Proteins (GFP) in Embryonen des Zebrafisches. In dem von ihnen etablierten Testsystem gibt sich ein regulatorischer DNA-Abschnitt dadurch zu erkennen, dass der Embryo GFP produziert und damit grün fluoresziert. In 17 Fällen erhielten McCallion et al. fluoreszierende Zebrafisch-Embryonen.
Mit den fünf gebräuchlichsten Programmen zur Genomanalyse konnten nur zwischen 29 und 61% der tatsächlich gefundenen regulatorisch wirksamen DNA-Sequenzen erkannt und prognostiziert werden. Damit ist gezeigt, dass die bisher verfügbaren Computerprogramme weiterentwickelt werden und auf eine solidere experimentelle Basis gestellt werden müssen. Diese Resultate unterstreichen auch den großen Bedarf an weiteren methodischen Zugängen, um bedeutsame DNA-Bereiche zu identifizieren. Durch Anwendung der bereits verfügbaren Techniken auf weitere Genbereiche ist ein erheblicher Kenntniszuwachs über weitere funktionelle Sequenzen zu erwarten.
Sollte ein erheblicher Anteil der im Genom niedergelegten Information bisher unerkannt sein, hätte das auch weitreichende Konsequenzen für die Anwendung molekularbiologischer Daten für die Analyse molekularer Stammbäume. Dann müsste – auch darauf deuten die Autoren hin – bei der Interpretation molekularbiologischer Daten im Sinne phylogenetischer Fragestellungen im Lichte dieser Erkenntnisse sehr viel sorgfältiger vorgegangen werden.
[McGaughey DM, Vinton RM, Huynh J, Al-Saif A, Beer MA & McCallion AS (2007) Metrics of sequence constraint overlook regulatory sequences in an exhaustive analysis at phox2b. Genome Res. (download advance articles 10.12.2007): http://www.genome.org/cgi/reprint/gr.6929408v1]
|
|
Die Polarregionen bieten unwirtliche Lebensbedingungen und sind daher artenarm. Infolge der Klimaerwärmung und den daraus resultierenden besseren Lebensbedingungen wird die Einwanderung von neuen Pflanzen- und Tierarten erwartet. Welche Faktoren bestimmen die Einwanderung?
Die Inselgruppe um Spitzbergen (= Svalbard-Archipel) eignet sich gut für die Untersuchung der Geschwindigkeit von Migrationsvorgängen (Einwanderungen), da sie während der letzten Eiszeit vor 20 000 Jahren konventioneller Zeitrechnung vollständig von Eis bedeckt war, und sehr wahrscheinlich die gesamte Flora seitdem neu einwanderte. Svalbard liegt nördlich des Polarkreises und weit entfernt von potentiellen Ausgangsregionen der Wiederbesiedlung (Grönland, Nordrussland, Nordskandinavien). Seine Tundrenvegetation zählt nur etwa 130 verschiedene Blütenpflanzenarten. Eine Forschergruppe (Alsos et al. 2007) sammelte 4439 Exemplare von neun arktischen Blütenpflanzenarten sowohl auf Svalbard als auch in den genannten möglichen Quellregionen. Die neun Arten aus sechs Familien wurden so ausgewählt, dass sie hinsichtlich ihrer ökologischen Ansprüche für die Flora Svalbards repräsentativ sind. Drei Arten haben gefiederte Ausbreitungseinheiten (Samen bzw. Früchte) für die Windverbreitung, weitere drei fleischige Früchte für die Verbreitung durch Vögel und Säugetiere und die restlichen drei sind hinsichtlich der Verbreitung nicht spezialisiert.
Durch Vergleich des „genetischen Fingerabdruckes“ aller 4439 Exemplare wurde für jede Art ermittelt, aus welcher Ausgangsregion sie den Svalbard-Archipel kolonisierte. Außerdem wurde abgeschätzt, wieviele verschiedene Samen jeder Art mindestens nach Svalbard gelangt sein mussten, um die dort vorgefundene genetische Vielfalt zu erklären.
Die hauptsächliche Besiedlung durch die untersuchten neun Arten nach der Eiszeit erfolgte aus Nordrussland, mit Abstand gefolgt von Nordskandinavien und Grönland. Das dürfte damit zusammenhängen, dass zwischen Svalbard und Nordrussland im Gegensatz zu Skandinavien im Winter eine Verbindung durch verdriftende Eisschollen besteht, mit deren Hilfe die Ausbreitungseinheiten gut transportiert werden können. Bei mehreren Arten waren die in Svalbard nachgewiesenen Populationen unterschiedlich und konnten entsprechend auch unterschiedlichen Herkünften zugewiesen werden. Beispielsweise gehen die Vorkommen der Rauschbeere (Vaccinium uliginosum) sowohl auf russische, als auch grönländische Quellen zurück, während etwa die Krähenbeere (Empetrum nigrum) nur aus Grönland und die Moltebeere (Rubus chamaemorus) nur aus Nordrussland stammen. Für die Alpen-Gänsekresse (Arabis alpina) konnte keine Entscheidung getroffen werden, da diese Art in den fraglichen Gebieten genetisch einheitlich ist.
Bei den übrigen Arten wurde abgeschätzt, dass jeweils mehrere Besiedlungen stattgefunden haben mussten, indem im Falle der Moltebeere mindestens 6, bei der Silberwurz (Dryas octopetala) sogar 38 Ausbreitungseinheiten unabhängig voneinander zu den Inseln Svalbards gelangten und sich dort erfolgeich etablieren konnten. Wegen der anzunehmenden fehlgeschlagenen Versuche dürften in Wirklichkeit viel mehr Ausbreitungseinheiten nach Svalbard gelangt sein.
Bei 6 Arten legte der genetische Befund neben der hauptsächlichen Quellregion noch eine zweite Ausgangsregion nahe: Für die vier kälteresistentesten Arten D. octopetala, S. herbacea, C. tetragona und S. rivularis sowie für die beiden am meisten wärmeliebenden (und folglich seltensten) Arten B. und V. uliginosum.
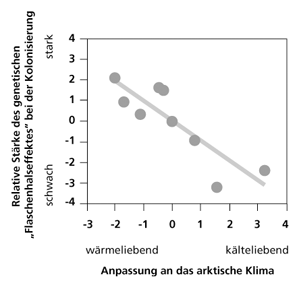 |
| Abb.1: Zusammenhang zwischen der Anpassung an das Klima und der genetischen Vielfalt bei den Pflanzen auf Svalbard. Nach Alsos et al. (2007) |
|
Die genetische Variabilität der untersuchten Pflanzen auf Svalbard korrelierte negativ mit ihrer Anpassung an das gegenwärtige dortige Klima: je besser eine Pflanzenart das Klima vertrug, desto größer war ihre genetische Vielfalt und umgekehrt (Abb. 1).
Nur von Juni bis August überschreiten die Mitteltemperaturen den Gefrierpunkt mit einem Maximum von 4,4 °C im Juli, und die Küstenregionen sind nur im Sommer für 6 Wochen eisfrei. Die Vegetation wird ermöglicht durch den Golfstrom, der während der 4-monatigen Polarnacht gegenüber dem auf dieser Breite üblichen Klima für relativ milde Winter sorgt.
Die Autoren kommen zu dem Schluss, dass bestimmend für die Neubesiedlung die ökologischen Bedingungen am Zielort sind und nicht die (offenbar immer in genügender Menge erfolgende) Verdriftung der Samen (durch Wind, Vögel oder z.B. Treibholz und -eis und selbst durch Säugetiere). Dieser Schluss wird unterstützt durch den Befund, dass 80 bis 90% der kälteresistenten Pflanzen in den potentiellen Ausgangsregionen auch auf Svalbard präsent sind, aber nur 40 bis 60% der relativ wärmeliebenden. Und damit würde ein wärmeres Klima auf Svalbard auch eine größere pflanzliche Vielfalt infolge erfolgreicher Migration erwarten lassen. Die gute Umweltresistenz bestimmter Pflanzensamen ist bekannt (Brown 2001, vgl. Lindemann 2001) und es können sich Pflanzen binnen weniger Jahre nicht nur vom Festland auf Inseln ausbreiten, sondern sich sogar mikroevolutiv an das neue Habitat anpassen (Cody et al. 1996, vgl. Kutzelnigg 1997). Auf der Basis der konventionellen Zeitrechnung veranschlagen Alsos et al. im Online support (www.sciencemag.org/cgi/content/full/316/5831/1606/DC1) 10 000 Jahre seit Beginn der Wiederbesiedlung. Aber auch wenn nur einige Prozent der verdrifteten Samen sich erfolgreich ansiedeln könnten, so wären selbst für Dryas octopetala nur einige 1000 Samen nötig. Die Leistungsfähigkeit der Transportprozesse dürfte die von Alsos et al. geschätzte Verdriftung von etwa einer Ausbreitungseinheit alle 2 Jahre bei weitem übersteigen (was einer erfolgreichen Ansiedlung einer Pflanze nur einmal pro Vierteljahrtausend entspräche). Damit stellt diese Studie indirekt die Datierung der letzten Eiszeit in Frage: Wäre Svalbard schon so lange teilweise eisfrei, müsste eine höhere genetische Vielfalt bestehen.
Nach einer größeren Katastrophe wäre jedenfalls zu erwarten, dass verwüstete, aber klimatisch günstige Regionen von vielleicht nur wenigen Ausgangspunkten aus auch über große Entfernungen rasch – innerhalb von Jahren – und durch nur wenige – einige Dutzend – Exemplare jeder Art wiederbesiedelt werden können, und genau das erwartet und sieht man ja auch heute bei der Artenmigration infolge der globalen Erwärmung.
[Alsos IG, Eidesen PB, Ehrich D, Skrede I, Westergaard K, Jacobsen GH, Landvik JY, Taberlet P & Brochmann C (2007) Frequent long-distance plant colonization in the changing arctic, Science 316, 1606-1609; Brown K (2001) Patience yields secrets of seed longevity, Science 291, 1884-1885; Cody ML & Overton JM (1996) Short-term evolution of reduced dispersal in island plant populations. J. Ecology 84, 53-61; Kutzelnigg H (1997) Schnelle Evolution der Samenverbreitung bei Pflanzenpopulationen auf küstennahen Inseln. Stud. Int. J. 4, 39-40; Lindemann WB (2001) Langlebige Pflanzensamen. Stud. Int. J. 8, 101.]
|
|  |

