|
|
|
|
Nachdem im 19. Jahrhundert im südwestdeutschen Sandsteinkeuper mehrere bruchstückhafte Schildkrötenreste gefunden worden waren, entdeckte man nach der Jahrhundertwende gut erhaltene Exemplare. Heute kennt man über ein Dutzend Schildkröten aus der europäischen Obertrias, die mit einer Ausnahme alle aus Württemberg stammen. Es handelt sich um die geologisch ältesten Schildkröten der Erde. Ihre Herkunft ist unbekannt (Ziegler 1986, 136f.). Man unterscheidet zwei Gattungen. Die etwas jüngere Proganochelys aus dem Mittleren Stubensandstein und dem überlagernden Knollenmergel (Mittlerer Keuper, Obere Trias) hat einen flachen Panzer und gilt als Wasserbewohner. Sie trug Zähne auf den Gaumenknochen und über den Hals- und Schwanzwirbeln waren (zum Schutz?) Knochendornen ausgebildet, die ein Zurückziehen von Kopf, Hals und Schwanz unter den Panzer verhinderten. Diese Merkmale kennzeichnen sie nach dem Stuttgarter Wirbeltierpaläontologen R. Wild (1998) als die am ursprünglichsten gebaute Schildkröte überhaupt (vgl. Müller 1985, 73f.). Im Unteren Stubensandstein, genauer: im ersten Stubensandstein-Horizont des Welzheimer und Murrhardter Waldes (nordöstlich Stuttgart) fand man die noch ältere Proterochersis (Hagdorn & Simon 1988, 85f.; vgl. Bachmann & Brunner 1998, 254). Auch wenn inzwischen neuere Schildkrötenfunde aus der Obertrias von Grönland, Argentinien, Südafrika und Thailand bekannt wurden, ist nach Wild Proterochersis immer noch die geologisch älteste Schildkröte der Erde (vgl. Benton 1993, 683f.). Ihr hochgewölbter Panzer kennzeichne sie als Landbewohner. Wie Wild (1998, 64) kürzlich in einem zusammenfassenden Beitrag über die südwestdeutschen Keuper-Vierfüßer schrieb, gehörte Proterochersis wahrscheinlich zu den Halswender-Schildkröten. Diese heutige Unterordnung zieht mit Hilfe eines anderen Mechanismus den Kopf unter den Panzer zurück als die heutigen Halsberger-Schildkröten (vgl. Kuhn-Schnyder & Rieber 1984, 214f.). Die knappe Beschreibung Wilds läßt immerhin erkennen, daß die ältere Proterochersis deutlich "moderner" gebaut war als die etwas jüngere Proganochelys. Möglicherweise ist dieser Befund eine Stütze für die inzwischen mehrfach belegte These des amerikanischen Paläontologen S.J. Gould (1998, 239-261), wonach in der Geschichte des Lebens keine Zunahme an Komplexität festzustellen ist, wenn die fossil nacheinander auftretenden Vertreter von Organismengruppen methodisch korrekt, nämlich in streng quantifizierbarer Weise vergleichend analysiert werden. Die gut erhaltenen Keuper-Schildkröten sind v.a. im Museum am Löwentor des Staatlichen Museums für Naturkunde in Stuttgart ausgestellt. [Bachmann GH & Brunner H (1998) Nordwürttemberg. Sammlg. Geol. Führer 90. Berlin, Stuttgart; Benton MJ (1993) Reptilia. In: Benton MJ (ed.) The Fossil Record 2, London, S. 681-715; Gould SJ (1998) Illusion Fortschritt. Frankfurt/M.; Hagdorn H & Simon T (1988) Geologie und Landschaft des Hohenloher Landes. Sigmaringen; Kuhn-Schnyder E & Rieber H (1984) Paläozoologie. Stuttgart, New York; Müller AH (1985) Lehrbuch der Paläozoologie, Bd. III/2. Jena; Wild R (1998) Zwischen Land und Meer. Saurier der Keuperzeit. In: Heizmann EPJ (Hg.) Vom Schwarzwald zum Ries, München, S. 57-66; Ziegler B (1986) Der schwäbische Lindwurm. Stuttgart.] MS |
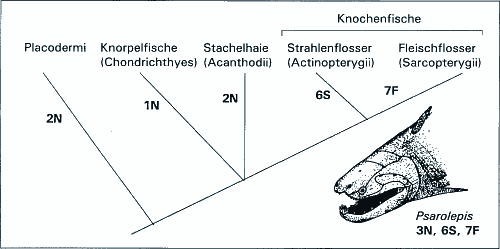
Der Ursprung der Fische wird durch Fossilfunde nicht erhellt. Die Fische werden in die Knorpelfische (Haie u.a.), Stachelhaie (Acanthodii) und Knochenfische unterteilt, letztere in die große Gruppe der Strahlenflosser (Actinopterygii) und die Fleischflosser (Sarcopterygii). Von den Fleischflossern werden die Landwirbeltiere (Tetrapoda, Vierbeiner) abgeleitet. Verfolgt man den Fossilbericht in die Vergangenheit zurück, werden die Fleischflosser vielfältiger, bis im Devon eine große Bandbreite von ihnen neben wenigen Strahlenflossern angetroffen wird; dann aber bricht der Fossilbericht abrupt ab (Ahlberg 1999). Aus dem Silur sind nur isolierte Schuppen und kleine Bruchstücke von Knochenfischen bekannt. Auch phylogenetische Analysen, die auf molekularen oder morphologischen Daten basieren, konnten nicht klären, welches der letzte gemeinsame Vorfahre der Fleischflosser oder der Knochenfische als Ganzes gewesen sein könnte. Von der Grenze Silur/Devon wird nun ein neuer Fund gemeldet. Zhou et al. (1999) entdeckten in Yunnan/Südchina die Gattung Psarolepis als nunmehr ältesten Knochenfisch. Doch wie so oft weist die neue Gattung ein Merkmalsmosaik auf, das sie als evolutionäre Übergangsform mehr als fragwürdig erscheinen läßt. Ahlberg (1999) spricht von einem unerwarteten Mix von Eigenschaften, der eine Neubewertung phylogenetischer Vorstellungen von großen Teilen des Wirbeltierstammbaums erforderlich mache. Schädelkapsel, Zähne und weitere Merkmale gleichen Fleischflossern. Die zähnetragenden Knochen der Schnauze und die Wangenknochen entsprechen denen der Strahlenflosser. Der Schultergürtel dagegen ähnelt allen Knochenfischen nicht. Ein großer Stachel ragt direkt vor der Brustflosse heraus - eine Situation, die den ausgestorbenen Placodermen entspricht. Andererseits scheint Psarolepis auch Stacheln der Spitze der medianen Flossen zu besitzen, was wiederum an die Stachelhaie erinnert (vgl. Abb. 1). |
|
|
Aufgrund dieses Merkmalsmosaiks ist die phylogenetische Position von Psarolepis nicht bestimmbar. Eine evolutionäre Übergangsform sollte zudem nicht ein Merkmalsmosaik vieler verschiedner Gruppen aufweisen, sondern im Gegenteil in möglichst vielen Merkmalen unspezialisiert sein. Dies ist Psarolepis gerade nicht, so daß durch diesen Fund hypothetische evolutionäre Zusammenhänge der Fischgruppen verwirrender und nicht klarer werden. [Ahlberg PE (1999) Something fishy in the family tree. Nature 397, 564-565; Zhou M, Yu X & Janvier P (1999) A primitive fossil fish sheds light on the origin of bony fishes. Nature 397, 607-610.] RJ |
Ascomyceten (Schlauchpilze) - dazu gehören Mutterkornpilz, Hefen, Pinselschimmel etc. - und die Basidiomyceten (Ständerpilze) bilden taxonomisch die höheren Pilze (Eumyceten) (Schlegel 1976). Aus der berühmten paläobotanischen Fundstätte in Rhynie, Aberdeenshire, Schottland aus dem Unteren Devon beschreiben Taylor, Hass und Kerp (1997) Fossilmaterial von Ascomyceten. Sie fanden Fruchtkörper (Perithezien) der Pilze zusammen mit aufrechten Stämmen und Wurzeln von frühen Landpflanzen der Gattung Asteroxylon, einer bärlappähnlichen krautigen Pflanze. Verschiedene Stadien aus dem Lebenszyklus der Pilze konnten fossil nachgewiesen (nämlich Perithezium, Asci [die namensgebenden schlauchartigen Gebilde] und Ascosporen) und so die Zuordnung zu den Ascomyceten abgesichert werden. Die Autoren verweisen darauf, daß bereits aus dem Silur Fossilien vorliegen, die als Ascomyceten interpretiert worden sind, diese Zuordnung allerdings umstritten sei und in der Literatur alternative Erklärungen diskutiert werden. Die nunmehr fossil nachgewiesenen Strukturen sind jedoch nach Aussagen der Autoren morphologisch identisch mit modernen Pyrenomyceten. Damit behaupten die Autoren nachgewiesen zu haben, daß die Pyrenomyceten entgegen den Ergebnissen mancher molekularer Pilz-Stammbaum-Rekonstruktionen innerhalb der Ascomyceten bereits lange Zeit als eigenständige Linie etabliert sind. Sie erhoffen sich mit diesen neuen Daten die Möglichkeit, molekularbiologische Befunde, die zur Konstruktion von molekularen Uhren herangezogen werden, zu kalibrieren. Paläontologische Befunde machen also eine Korrektur der konstruierten phylogenetischen Aufspaltung erforderlich und zwar in der Weise, daß die Aufspaltung zeitlich immer weiter in die Vergangenheit zurück verschoben werden muß. [Schlegel HG (1976) Allgemeine Mikrobiologie. 4. Aufl., Stuttgart; Taylor TN, Hass H & Kerp H (1997) A cyanolichen from the Lower Devonian Rhynie Chert. Am. J. Bot. 84, 992-1004; Taylor TN, Hass H & Kerp H (1999) The oldest fossil ascomycetes. Nature 399, 648.] HB |
Verschiedene Gattungen von Tintenfischen (Familie Sepiolidae) haben Leuchtdrüsen auf ihrem Tintenbeutel, die lumineszierende Bakterien enthalten. Bei Gefahr können die Bakterien von den Tintenfischen teilweise als leuchtende Wolke ausgestoßen werden. Die Tintenfisch-Gattung Euprymna besitzt außerdem auch mit Leuchtbakterien gefüllte Organe mit Linsen und Reflektoren. Bei vielen Symbiose-Partnern besteht eine Übereinstimmung in den Verwandtschaftsverhältnissen, die gewöhnlich als parallele Evolution gedeutet wird. Sind die Wirte nah miteinander verwandt, so sind es meistens auch die Gäste. Nishiguchi (1998) und Mitarbeiter untersuchten diesen Zusammenhang in der Tintenfisch-Leuchtbakterien-Symbiose. Bestimmte Sequenzabschnitte aus dem Erbgut beider Organismen wurden miteinander verglichen. Es zeigte sich, daß die Ähnlichkeitsbäume beider Gruppen (Tintenfisch vs. Bakterien) übereinstimmen. Einen vergleichbaren Befund gibt es z.B. auch von Ameisen-Pilz-Symbiosen. Hier ist der übereinstimmende Stammbaum leicht zu erklären. Neue Königinnen nehmen ein Stück des benötigten Pilzes aus ihrem alten Nest mit. Somit bleiben die Partner zusammen und sind einer ähnlichen Mikroevolution unterworfen. Bei der Tintenfisch-Leuchtbakterien-Symbiose müssen sich aber die Partner immer wieder neu finden, denn junge, frisch geschlüpfte Tintenfische besitzen noch keine symbiontischen Bakterien. Sie nehmen diese aus dem Meerwasser in ihre speziellen Organe auf. Interessanterweise finden sich aber bei den Tintenfischen und Leuchtbakterien die jeweils co-evolvierten Partner immer wieder (neu) zusammen. Setzt man experimentell junge Euprymna-Tintenfische Leuchtbaktieren (Vibrio-Arten von verschiedenen anderen Tintenfischen) aus, so wird Euprymna am besten von solchen Bakterien besiedelt, die ursprünglich von der eigenen Art stammten. Damit scheint die Präferenz für den jeweiligen Symbiosepartner genetisch festgelegt zu sein. Co-Evolution, die in einigen Fällen auch eine parallele Evolution darstellt, findet sich in der Natur häufiger. Beispielsweise sind bestimmte Orchideen auf spezielle Bienen angewiesen, die wiederum Duftstoffe genau dieser Orchideen für die Anlockung ihrer Weibchen benötigen. Der symbiontische Pilz der Blattschneiderameisen kann nur in der Obhut der Ameisen überleben und die Ameisen nur mit Pilz. Vielfach wird theoretisch davon ausgegangen, daß die Partner ursprünglich in einer losen Gemeinschaft zusammenlebten (eventuell sogar ein Partner den anderen parasitierte) und dann im Laufe der Zeit durch eine wechselseitige Anpassung in eine Symbiose eintraten. Nicht selten wird aber die Existenz einer Symbiose erst dann erklärbar, wenn beide Partner ihre spezifischen Eigenschaften schon besitzen und einbringen. Daß Symbiose-Partner eine parallele Mikroevolution durchlaufen, ist nicht überraschend, jedoch bleibt der Anfang einer solchen Partnerschaft meist offen. [Nishigushi MK, Ruby EG & McFall-Ngai MJ, Applied Environmental Microbiology 64, 3209-3213] KN |
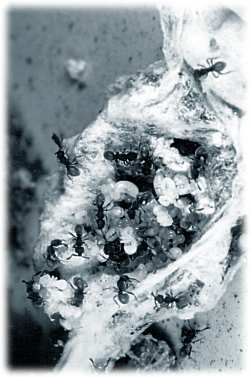
Ameisen aus der Gruppe der Stachelameisen (Ponerinen) haben keine claustrale Koloniegründung und gelten deshalb als primitiv, es gibt aber eine Stachelameise Discothyrea oculata aus Kamerun, bei der die Königin trotzdem die Gefahren einer eigenen Jagd umgeht. Die Königinnen haben eine ganz ausgefallene Kinderstube entdeckt: Sie nisten sich in von Seide umsponnenen Eiballen kribellater Spinnen (Ariadna spec.) ein. Kribellate Spinnen produzieren keine klebrigen Seidenfäden zum Beutefang, sondern umspinnen die Haltefäden des Netzes mit feinsten "Fusseln" aus Seide. Insekten bleiben dann mit ihren Körperteilen und Borsten in diesen "Fusseln" hängen. In den Eiballen dieser Spinnen haben die Ameisen-Königinnen nun Nahrung durch Spinneneier und Schutz durch die Seidenhülle. Die Königin braucht ihre Unterkunft nicht verlassen und ist gleichzeitig auch nicht auf eigene Vorräte angewiesen. Experimente zeigten, daß die Ameisen in der Lage sind, die Seide zu manipulieren und damit das Innere des Eiballen nach ihrem Gutdünken zu gestalten. Die ursprüngliche Koloniebildung dieser Ameisenart muß allerdings semi-claustral gewesen sein, denn wurden in Experimenten den Königinnen leere Seidenhüllen angeboten, so begaben sie sich auf Suche nach Spinneneiern, um damit Nahrung herbeizuschaffen. Es ist fraglich, ob man eine Art der Koloniegründung hochentwickelt (claustral) oder eine andere primitiv (semi-claustral) nennen kann. Jede Ameisenart hat ihre Besonderheiten bei der Koloniegründung und jede Art überlebt. Je nach Umwelt ist die eine oder andere Koloniegründung von Vor- oder Nachteil. Auch die vorgeblich primitiveren Stachelameisen stehen anderen Ameisen in der Komplexität ihrer Verhaltensweisen in nichts nach. [Dejean A & Dejean A (1998) Insectes sociaux 45, 343-346.] KN |
50 der 128 Arten der Familie Cuculidae (Kuckucke) brüten nicht selber, sondern legen ihre Eier als obligate Brutschmarotzer in fremde Nester. Eine große Zahl von Vogelarten dient dabei als Wirtsvögel. Seit langem ist bekannt, daß keine Kuckucksart nur eine einzige Wirtsvogelart wählt; die Wirtsvögel wechseln vielmehr räumlich und zeitlich. Weiterhin ist bekannt, daß die Eier von Kuckucksweibchen in Größe und Färbung den Eiern des jeweiligen Wirtsvogels sehr nahe kommen und dadurch die Wirte nicht veranlassen, das Ei als fremd zu erkennen und hinauszuwerfen oder ihr Gelege zu verlassen. Der in Deutschland heimische Cuculus canorus legt etwa 15 bis 20 verschiedene Typen von Eiern. Ein einzelnes Weibchen legt aber zeitlebens übereinstimmende Eier, nach Möglichkeit in die Nester seiner bevorzugten Wirtsvogelart. Wahrscheinlich sind Prägungsvorgänge für diese Präferenz verantwortlich. Falls einmal ein Weibchen "zufällig" ein Ei in ein Nest einer Vogelart legt, von der es nicht selber aufgezogen wurde, und dieses Ei von der neuen Art akzeptiert wird, so ist damit eine neue Wirtsvogelart erschlossen. In Nestern von 90 der 130 heimischen Singvogelarten wurden Kuckuckseier gefunden, die Zahl der Hauptwirte ist aber erheblich geringer (Meise 1969). Aus evolutionstheoretischer Sicht könnte erwartet werden, daß eine immer weitergehende Spezialisierung einzelner Kuckuckspopulationen auf bestimmte Wirte erfolgt, so daß zunehmend neue, spezialisierte Kuckucksarten entstehen. Ein solcher Vorgang wäre ein klassischer Fall von Mikroevolution. Trotz andernorts beobachteter rascher Artbildung (ein kurzer Überblick findet sich bei Junker & Scherer 1998, S. 290f.) im Rahmen mikroevolutiver Vorgänge ist dies aber bisher nicht eingetreten. Eine Erklärung für diesen Sachverhalt bietet erstmals eine Publikation einer amerikanisch-japanischen Forschergruppe (Marchetti et al. 1998): in einem Waldgebiet Japans wurden Blutproben von 162 adulten Vögeln (83 männlich, 79 weiblich) und 136 Küken der Art Cuculus canorus entnommen. Bei den Küken wurde vermerkt, welcher der drei Hauptwirte der Region sie beherbergte. Acht Kuckuck-spezifische DNA-Mikrosatellitenmarker wurden verwendet, um zu bestimmen, welche Eltern zu welchen der 136 Küken gehörten. 84% der Küken konnten wenigstens einem der insgesamt 162 adulten Vögel als Elterntier zugeordnet werden. Aus diesen Daten wurden Rückschlüsse auf Paarungsverhalten und Wirtsvogel-Präferenzen gezogen. Es ergab sich, daß Kuckucke beider Geschlechter vergleichbares polygames Paarungsverhalten zeigen: 7 von 15 Männchen und 3 von 18 Weibchen hatten mehr als einen Partner. Deutlicher unterschiedlich war die Auswahl der Wirtsvogelarten: 7 von 19 Männchen, aber nur 2 von 24 Weibchen hatten Nachkommen in mehr als einer der drei in dieser Region hauptsächlich benutzten Wirtsvogelarten. Der Unterschied zwischen Männchen und Weibchen ist signifikant (p<0.05). Nur 5% aller Eier wurden von Weibchen nicht in ein Nest der eigenen Wirtsvogelart gelegt. Männchen und Weibchen unterscheiden sich also hinsichtlich ihrer Wirtsvogelpräferenz, indem Weibchen spezialisierter sind als Männchen. Einigen Individuen konnten mehrere Nachkommen zugeordnet werden. Sie hatten sich mit einem Partner derselben oder einer anderen Wirtsvogelpräferenz verpaart. Da sich nicht nur Kuckucke mit derselben Wirtsvogelpräferenz verpaarten, sind keine guten Voraussetzungen für eine Rassen- oder Unterartbildung gegeben. Als Erklärung bietet sich nach Ansicht der Autoren an, daß Männchen und Weibchen ihren Fortpflanzungserfolg über verschiedene Strategien optimieren: die Männchen durch ein Maximum an Paarungen, die Weibchen durch an einen bestimmten Wirt bestangepaßte Eier, wodurch ein Maximum an überlebenden Jungvögeln erreicht wird. Der ständige Genfluß über die Männchen verhindert dabei jedoch eine Artaufspaltung. Über die genauen Mechanismen herrscht noch keine Klarheit. Die Autoren schlagen vor, daß die Vererbung der für die Morphologie der Eier verantwortlichen Gene nur zwischen Mutter und Tochter erfolgen soll, ein Postulat, das weitere Forschung verlangt (Morell 1998). [Junker R & Scherer S (1998) Evolution - ein kritisches Lehrbuch. Gießen; Marchetti K et al. (1998) Host-race formation in the common cuckoo. Science 282, 471-472; Meise W (1968) Kuckucksvögel. In: Grzimeks Tierleben, Enzyklopädie des Tierreiches, Bd 8 (Vögel 2), Zürich, S. 341-376; Morell V (1998) Male mating blocks new cuckoo species. Science 282, 393.] WL |
Schon manche berühmten Lehrbuchbeispiele sind späteren Überprüfungen zum Opfer gefallen. Nun ist offenbar auch eines der in evolutionstheoretischen Lehrbüchern meistzitierten Beispiele an der Reihe: Die Geschichte von der Anpassung und Selektion des Birkenspanners Biston betularia) muß neu geschrieben werden. Das geht jedenfalls aus Studien hervor, die Majerus 1998 in seinem Buch "Melanism: Evolution in Action" veröffentlichte. Nach bisheriger Lesart, die sich vor allem auf Arbeiten von Kettlewell (1973) stützte, nahmen zwischen 1850 und 1920 die dunklen Formen (forma carbonaria) des Falters im Zuge der Industrialisierung und der damit verbundenen Umweltverschmutzung erheblich zu und die hellen (forma typica) entsprechend ab. Nach 1950 drehte sich der Trend wieder um. Als Ursache für diese Verschiebungen der Häufigkeiten wurde das Ausmaß der Luftverschmutzung angenommen, aufgrund derer die hellen Flechten auf den Borken der Bäume abstarben, was wiederum dazu führte, daß die dort ruhenden hellen Formen des Birkenspanners nicht mehr gut getarnt waren und häufiger von Vögeln erbeutet wurden: Selektion in Aktion - so ist wohl jedem Biologen dieses Beispiel bekannt. Kettlewell hatte eine Reihe von Experimenten durchgeführt, um dieses Szenario empirisch zu belegen. Beispielsweise hatte er typica und carbonaria auf verschmutzte und unverschmutzte Borken gesetzt; später wurden dann mehr getarnte als auffällige Formen gefunden. In Laborstudien fand Kettlewell heraus, daß die beiden Formen denjenigen Untergrund bevorzugten, auf dem sie besser getarnt waren. Es gab schon länger Kritik an dieser Geschichte; Majerus (1998) faßte sie nun in zwei Kapiteln seines Buches zusammen. Der schwerwiegendste Einwand ist, daß sich die Birkenspanner nahezu niemals auf Baumstämmen niederlassen; nur zweimal wurde diese Position während 40 Jahren intensiver Forschung beobachtet. Die Experimente Kettlewells gingen also an der natürlichen Situation vorbei; die Ruhestellen des Birkenspanners sind in Wirklichkeit gar nicht bekannt. Eine weitere Schwächung der Plausibilität der bekannten Lesart der Birkenspanner-Geschichte ist die Tatsache, daß die hellen Formen wieder zunahmen, bevor die Flechten sich wieder auf den Borken ausgebreitet hatten (nach 1950, als die Luftverschutzung wieder abnahm). Schließlich konnten Kettlewells Verhaltensstudien nicht bestätigt werden: Die Falter tendierten nicht dazu, solche Untergründen zu wählen, die zu ihrer Farbe paßten. Majerus fand noch weitere Fehler in Kettlewells Arbeit. Was kann man aus den neuen Einsichten schließen? Klar ist, daß die einfache Version der "Selektion in Aktion" nicht stimmen kann. Selektion mag eine (wesentliche?) Rolle bei der Verschiebung der Häufigkeiten der Formen des Falters gespielt haben und es mag einen Zusammenhang zwischen Umweltverschmutzung und dieser Verschiebung geben. Wenn dem so sein solte, dann ist dieser Zusammenhang viel komplizierter als bisher angenommen und unverstanden. Coyne (1998, 603) stellt in einer Rezension des Buches von Majerus fest: "B. betularia shows the footprint of natural selection, but we have not yet seen the feet." Klar ist lediglich, daß es sich um einen Fall schneller Mikroevolution handelt, wobei jedoch nur eine Verschiebung von Allelhäufigkeiten vorliegt. [Majerus MEN (1998) Melanism: Evolution in action. Oxford; Coyne JA (1998) Not black and white. Nature 396, 35-36.] RJ |
Für die Zellteilung benötigen Bakterien ein Protein, welches als FtsZ bezeichnet wird. Es bildet während der Teilung einen Ring, mit dessen Hilfe eine neue Zwischenwand eingezogen wird. Im Laufe der Zellteilung wird der FtsZ-Ring immer kleiner. Er zieht dabei eine neue Zellwand hinter sich her und schnürt die Tochterzellen schließlich voneinander ab. Homologe von FtsZ hat man interessanterweise in allen untersuchten Bakterien einschließlich Mycoplasmen (einfache zellwandlose Bakterien), sowie in Archaea (einzellige Lebensformen, die aufgrund molekularbiologischer Befunde von den Eubakterien und Eukaryoten abgegrenzt wurden) und Chloroplasten entdeckt. Es zeigt sich also, daß dieses Protein in einem großen Teil des Organismenreiches eine wichtige Rolle in der Zellteilung spielt. Jedoch hat man in Mitochondrien kein solches Protein gefunden und der Mechanismus der Teilung ist dort noch unklar. Alle bislang gefundenen Homologe von FtsZ sind am Anfang ihrer Struktur (N-terminal) konserviert, das heißt, die Proteine aus verschiedenen Organismen stimmen im vorderen Teil in ihrer Aminosäure-Sequenz sehr gut überein. Das Ende (C-terminal) muß aber mit anderen Proteinen in Kontakt treten und diese sind je nach Organismus verschieden, dementsprechend ist der C-terminale Bereich sehr variabel. Evolutionstheoretisch wird vermutet, daß FtsZ ein Protein ist, das sehr früh in der Evolution entstand und aufgrund seiner Wichtigkeit von allen Prokaryoten beibehalten wurde. Chloroplasten werden als Nachkommen von einverleibten cyanobakterien-ähnlichen Vorfahren angesehen und enthalten dementsprechend FtsZ. Auch die Archaea sollen letztlich aus Prokaryoten hervorgegangen sein. Die Ausnahme der Mitochondrien, die kein FtsZ enthalten, aber ebenfalls bakterielle Vorläufer haben sollen, wirft jedoch noch Fragen auf. Handelt es sich bei FtsZ um ein während der Evolution konserviertes Protein? Etwas Vergleichbares fand man bei Augen von verschiedenen Organismen. Der Schalter für die Augenentwicklung ist bei Insekten (Fruchtfliege, Drosophila) derselbe wie bei Wirbeltieren (Maus, Mus). Erfüllt eine Struktur eine bestimmte Aufgabe am besten, so ist es nicht einsichtig, warum in verschiedenen Organismen verschiedene Strukturen auftreten sollten. Beispielsweise werden in Uhren Zahnräder benötigt, und ein Uhrmacher wird nicht versuchen, andere Teile als Zahnräder zu verwenden, denn Zahnräder sind eben für diese Funktion optimal. Insofern muß man gleiche Bauteile in Lebewesen nicht notwendigerweise damit erklären, daß die Lebewesen einen gemeinsamen Vorfahren hatten. Ausnahmen (Mitochondrien, s.o.) stellen die Evolutionslehre vor das Problem, daß entweder erklärt werden muß, wie ein Übergang von FtsZ-haltigen Zellen (Vorfahren von Mitochondrien) zu solchen erfolgte, die kein FtsZ mehr benötigen (Mitochondrien) oder aber warum ein ähnlicher, sehr komplexer Vorgang (Zellteilung) mehrfach unabhängig (konvergent) entstanden ist. [Lutkenhaus J (1998) Current Biology 8, R619-R621.] KN |
Die amerikanische Weltraumbehörde NASA hat im Mai 1998 am Ames Research Center in Mountain View, California ein neues virtuelles Institut für Astrobiologie eingerichtet (Lawler 1999a, b). Dazu hat eine Kommission aus 53 Bewerbern 11 Institutionen (s. Tab. 1) ausgewählt, die mittels Internet interdisziplinär kooperieren sollen. Die Teilnehmer sollen über ein neues, von der NASA entwickeltes Hochgeschwindigkeits-Computer-Netzwerk (´Next Generation Internet´) kommunizieren. Das Netzwerk soll im Rahmen dieses Instituts seine Leistungsfähigkeit unter Beweis stellen und Kosteneinsparungen ermöglichen. |
Motiviert durch Entdeckungen von außergewöhnlichen Lebewesen in extremen Lebensräumen, wie z.B. in heißen Tiefseeschloten oder in basaltischem Gestein mehrere hundert Meter unter der Erdoberfläche, sollen Fragen im Zusammenhang mit der Entstehung des Lebens neu beleuchtet werden. Auch die Interpretation von Untersuchungsergebnissen von Meteoritenmaterial oder spektroskopischen Hinweisen auf Wasservorkommen z.B. auf dem Jupitermond Europa nähren Spekulationen über das Auftreten von Lebewesen im Kosmos, die geprüft und weiter untersucht werden sollen. Die Projekte werden jeweils in Zeiträumen von fünf Jahren geplant. Für 1998 standen 4 Millionen Dollar zur Verfügung, für 1999 sind 9 Millionen Dollar veranschlagt und im Jahr 2000 sollen es 20 Millionen Dollar sein. Vertreter der NASA haben die Hoffnung geäußert, daß in späteren Jahren 100 Millionen Dollar pro Jahr zur Verfügung stehen. Inzwischen wurde bekannt, daß sich ein spanisches Institut dem Astrobiologie-Zentrum anschließt und die spanische Regierung sich mit 7 Millionen Dollar beteiligen will (Anonymous 1999). Es ist bemerkenswert, mit welch hohem Einsatz und Aufwand die ungelösten Fragen der Entstehung der Lebewesen in Angriff genommen werden und man darf auf die Ergebnisse gespannt sein, die durch diese enormen Anstrengungen in den nächsten Jahren erzielt werden. Abschließend sei jedoch die skeptische Frage erlaubt, ob die bisher vorliegenden Resultate früherer, intensiver Bemühungen im Themenbereich Ursprung des Lebens einen solchen finanziellen Aufwand wirklich rechtfertigen, oder ob man beim Einsatz und der Verteilung von Forschungsmitteln nicht sorgfältiger prüfen sollte. [Anonymous (1999) Spain joins search for extraterrestrial life. Nature 397; Lawler A (1998a) Ames tackles the riddle of life Science 279, 1840-1842; Lawler A (1998b) Astrobiology institute picks partners. Science 280, 1338; Vaas R (1999) Astrobiologie bei der NASA. Naturw. Rdsch. 52, 203-204; Informationen im Internet (s. Vaas 1999): http://astrobiology.arc.nasa.gov/; http://www.reston.com/astro/bio.web.html.] HB | |||||||||||||||||||||||||||||||||||||||||

|