|
|
|
|
|
Die meisten Fossilien sind nur sehr bruchstückhaft erhalten. Isolierte Organe werden daher als Formgattungen beschrieben, zum Beispiel als Gattungen für Zapfen, für sterile Zweige, für Blätter, für Sporophylle (Blätter mit Sporenbehältern [Sporangien]), für Sporen, für Stämme (mit unterschiedlichen Bezeichnungen für verschiedene Erhaltungszustände) und für unterirdische Organe. Evolutionstheoretische Betrachtungen sind aufgrund dieser Situation erheblich erschwert und oft kaum möglich. Dies gilt teilweise auch für die Lepidophyten, besonders für deren ältere Überreste aus dem Oberdevon und Unterkarbon, wo die Rekonstruktion kompletter Pflanzengestalten einschließlich der wichtigen unterirdischen Organe bisher nicht möglich war (Bateman et al. 1992, 546).
Rekonstruktionen der Stammesgeschichte werden durch einen weiteren Befund erschwert: Vollständiger erhaltene Gattungen zeigen nämlich, daß die einzelnen Organe in unterschiedlichsten Ausprägungen kombiniert sein können. Beispielsweise können Gattungen mit monosporangiaten Zapfen (nur ein Sporentyp in einem Zapfen) baumförmig oder scheinkrautig sein; dasselbe gilt ebenso für Gattungen mit bisporangiaten Zapfen. Das heißt: Von der Ausprägung eines bestimmten Organs kann nicht auf den Zustand eines anderen geschlossen werden. Hier begegnet uns das Phänomen der Heterobathmie (Kombination sog. primitiver und sog. fortschrittlicher Merkmale), das im Pflanzenreich häufig angetroffen wird. Diese Situation verbietet es, aus isolierten Pflanzenfragmenten Schlüsse über den gesamten Bauplan zu ziehen (vgl. Bateman et al. 1992, 550). |
Aussagen über phylogenetische Zusammenhänge können aus diesen Gründen oft nur in allgemeiner Form im Sinne von gröberen Tendenzen einzelner Strukturen formuliert werden, wohingegen die theoretisch denkbaren Abfolgen bei verschiedenen Strukturen in vielen Fällen nicht zusammenpassen (vgl. Junker 1996). |
|
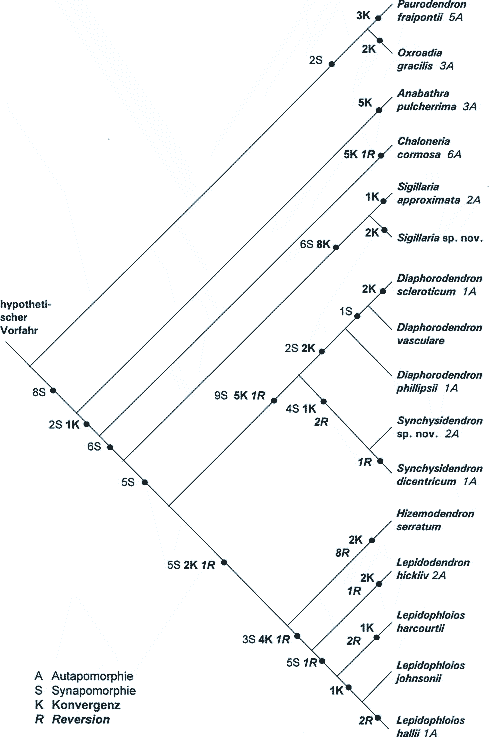
Aus dem Oberkarbon ist jedoch eine ganze Reihe insgesamt gut erhaltener Lepidophyten-Gattungen bekannt (Abb. 1), so daß aussagekräftige phylogenetische Analysen möglich sind. Viele oberkarbonische Arten sind auch aus dem Unterkarbon bekannt, kommen dort aber in der Regel in weniger gut erhaltener Form vor. Bateman et al. (1992) führten mit 16 Arten eine umfangreiche cladistische Analyse durch. Darin werden 69 vegetative und 46 generative Merkmale einbezogen. Abb. 2 gibt den "bevorzugten sparsamsten Baum" wieder (d. h. die Zahl der anzunehmenden Konvergenzen und Revisionen ist minimal; s. u.). Da in diesem Überblicksartikel auf die meisten Details nicht eingegangen werden kann, wurde im Dendrogramm lediglich angegeben, wieviele Synapomorphien, Konvergenzen und Reversionen jeweils auftreten (zu den Begriffen siehe Glossar). Dabei sind die einzelnen Merkmale allerdings von unterschiedlichem Gewicht. Einige wichtige Beobachtungen sollen im folgenden hervorgehoben und kommentiert werden. Der an Details interessierte Leser sei an die zugrundeliegende ausführliche Arbeit von Bateman et al. (1992) verwiesen. |
|
Konvergenzen und Reversionen.
Abgrenzung von Gattungen und Arten.Die in der cladistischen Analyse von Bateman et al. (1992) untersuchten Gattungen sind ziemlich gut abgegrenzt; sie unterscheiden sich voneinander durch jeweils eine beträchtliche Anzahl von Merkmalen. So stellen Phillips & DiMichele (1992, 571) beispielsweise fest, daß es auffällig verschiedene Megasporangium-Sporophyll-Komplexe gibt. Die Mikrosporangien und besonders die Mikrosporen ermöglichen scharfe Unterscheidungen zwischen Sigillaria, Diaphorodendron und den Gattungen mit Sporengattung Lycospora (Phillips & DiMichele 1992, 571).1 Innerhalb der Gattungen ist eine Auflösung in die verschiedenen Arten kaum durch qualitative Unterschiede möglich (Bateman et al. 1992, 526, Phillips & DiMichele 1992, 572). Möglicherweise lassen sich die Gattungen jeweils als Grundtypen (im Sinne von Scherer 1993) abgrenzen (siehe aber auch die Überlegungen im Schlußabschnitt).
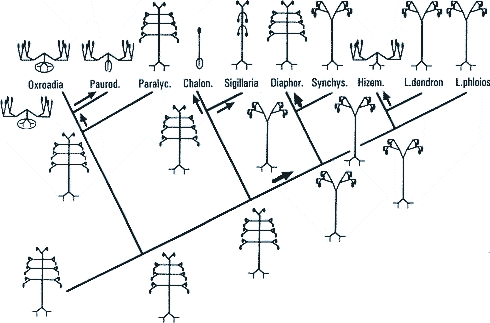
Teilanalysen.Bateman et al. (1992) haben gesonderte Dendrogramme unter Zugrundelegung nur der vegetativen bzw. nur der generativen Merkmale aufgestellt (Abb. 4 und 5). Dabei zeigen sich einige deutliche Unterschiede, die einleitend bereits angedeutet wurden: Manche Gattungen sind nach vegetativen Merkmalen primitiv, nach generativen jedoch fortschrittlich und umgekehrt. Vegetative und generative Trends sind nicht in Deckung zu bringen. Entweder die Zapfenmorphologie oder die Wuchsform ist als konvergent zu betrachten. Dies wird deutlich bei der scheinkrautigen Gattung Hizemodendron, die in der Zapfenmorphologie fortschrittlich, in der Wuchsform jedoch primitiv ist. Der umgekehrte Fall liegt bei Anabathra vor. Sigillaria ist insgesamt nach vegetativen Merkmalen fortschrittlicher als nach generativen, bezüglich der fünf in der Untersuchung berücksichtigten Blattpolstermerkmale jedoch wiederum primitiv. Die nach vegetativen Merkmalen insgesamt primitiveren Gattungen Diaphorodendron und Synchysidendron sind in allen diesen Blattpolstermerkmalen dagegen fortschrittlich (Bateman et al. 1992, 514). Zu beachten ist, daß auch bei Beschränkung auf den jeweiligen Teil des gesamten Merkmalsspektrums immer noch zahlreiche Konvergenzen und Reversionen zu verzeichnen sind. |
Die große Zahl an Konvergenzen und Reversionen ist evolutionstheoretisch gesehen problematisch, insbesondere bei wiederholtem Auftreten und je komplexer die betreffenden Merkmale sind. Bateman et al. (1992) und Bateman & DiMichele (1994) diskutieren eine Lösung, die auf eine Verkleinerung des Problems setzt. Vielleicht können größere morphologische Änderungen auf geringe genetische Unterschiede zurückgeführt werden, wie dies etwa von der Fruchtfliege Drosophila bekannt ist, wo homöotische Mutationen (bei denen ganze Bauteile an falschen Körperstellen realisiert werden) zu bedeutsamen Änderungen führen können. Solche wiederholt auftretende sprunghafte Evolution (Saltation) soll nach Bateman & DiMichele (1994) die Merkmalswidersprüche in den Cladogrammen verständlich machen. Dabei definieren diese Autoren "sprunghafte Evolution" als genetische Veränderung innerhalb einer Generation, die sich phänotypisch deutlich manifestiert und zu einer neuen evolutionären Linie führt (S. 62, 66). Die Richtung der Veränderung ist unerheblich (Vergrößerung oder - was häufiger ist - Verringerung der Komplexität). Die Autoren sprechen von einem "Neo-Goldschmidtschen Paradigma". Nach Ansicht von Bateman et al. (1992, 545), Bateman & DiMichele (1994) und Bateman (1994, 572ff.) könnte die chaotische Anordnung der Wuchsformen der Lepidophyten im sparsamsten Cladogramm auf Mutationen (möglicherweise eine einzige) in regulatorischen Genbezirken zurückzuführen sein, die ähnliche sprunghafte Änderungen bewirken.2 Solche Mutationen sollen in frühen ontogenetischen Stadien wirksam sein und dadurch weitreichende Änderungen zur Folge haben. Als Ausgangsform für die verschiedenen Wuchsformen käme ein baumförmiger Vorläufer in Betracht, der durch Heterochronie durch den Verlust eines Bauplanmoduls in andere, einfachere Wuchsformen überführt wurde (Bateman et al. 1992, 545; Bateman 1992, 88; Bateman & DiMichele 1991, 203). Davon durfte freilich die Ausprägung anderer Merkmale, z. B. der Zapfen nicht betroffen sein (vgl. Bateman 1992, 89). Mit diesem Konzept könnte evolutionstheoretisch ein Teil der Konvergenzen und vor allem der Reversionen erklärt werden. Allerdings läuft diese Lösung darauf hinaus, als Vorläuferform eine ziemlich "komplett" ausgestattete, mit allen vier Bauplanmodulen versehene Vorläuferform zu postulieren (Bateman 1994, 568). Die Entstehung der Merkmale einer solchen hypothetischen Ausgangsform wäre mit diesem Szenario nicht zu begründen. Die Konvergenzen in der Wuchsform entstünden durch unterschiedliche Variation oder Wegfall einzelner Bauteile (Bateman & DiMichele 1994, 83). Diese Vorgänge sind also nicht geeignet, die Entstehung des evolutionär Neuen verständlich zu machen. Das Auftreten von Mutationen in regulatorischen Bezirken alleine genügt zudem nicht; die Mutanten müssen einen Weg zum Überleben finden. Aufgrund eines extremen genetischen Flaschenhalses wird die genetische Vielfalt im Vergleich zur Elternpopulation stark reduziert, was zu anfangs verringerter Konkurrenzfähigkeit führt. Eine Regulationsmutation bedeutet nämlich das Verlassen eines adaptiven Optimums. "Experimente" sprunghafter Evolution werden daher in der Regel fehlschlagen, und nur gelegentlich zum Erfolg führen, nämlich wenn auch die Umweltbedingungen es erlauben (vgl. Bateman & DiMichele 1994, 81, 84): Es wird postuliert, daß solche Sprünge in einer Art ökologischem Vakuum, d. h. auf Pionierstandorten mit wenig Konkurrenz erfolgen (Bateman & DiMichele 1994, 83; Bateman & DiMichele 1991, 203). Das geschilderte Überlebensszenario wäre also dann plausibel, wenn zum einen geologisch entsprechende paläoökologische Bedingungen nachweisbar sind und wenn zum anderen die postulierten genetischen Mechanismen sich experimentell bestätigen lassen. Auf diese beiden Aspekte wird hier nicht eingegangen (vgl. dazu Bateman & DiMichele 1994, 83f. und dort angegebene weiterführende Literatur). Eine wichtige Konsequenz der saltatorischen Evolution, die in der skizzierten Weise erfolgt, ist die Veränderung von weiteren Merkmalen, die sich quasi automatisch als Folge einer Regulationsmutation weiterhin einstellen. Wird z. B. die Morphogenese (Formbildung) durch Heterochronie verkürzt, dann wird nicht nur ein bestimmtes Bauplanmodul wegfallen, sondern es werden konsequenterweise damit gekoppelte Merkmale verschwinden oder verändert werden. Beim Extrembeispiel Hizemodendron (s. Abb. 1) ist allerdings nicht ersichtlich, daß die Häufigkeit der in dieser Gattung zu postulierenden Konvergenzen und vor allem der Reversionen im Zusammenhang mit der postulierten Heterochronie stehen. Auch die konvergenten Merkmale beim Paar Diaphorodendron / Synchysidendron und bei Sigillaria dürften weithin ungekoppelt sein; bei Sigillaria sind Merkmale des Habitus, der Stigmarien, des Cortex, des Periderms, des Blattpolsters, der Blätter, des Sporophylls sowie der Mega- und Mikrosporen betroffen. Das evolutionstheoretische Problem der häufigen Reversionen erscheint also durch die Annahme einer schnellen heterochronischen Reduktion nicht wesentlich entschärft zu werden. |
Bateman (1994) diskutiert zwei Möglichkeiten für die Wuchsform des hypothetischen Vorfahren der in der cladistischen Analyse von Bateman et al. (1992) berücksichtigten Gattungen, nämlich eine Ausprägung wie bei der scheinkrautigen Oxroadia oder wie bei der baumförmigen Gattung Paralycopodites (= Anabathra). In beiden Fällen muß die gleiche Anzahl an konvergenten Übergängen in der Architektur angenommen werden. Bateman (1994, 568) bevorzugt Paralycopodites als geeignetste Außengruppe. Das würde bedeuten, daß die ursprünglichste Ausprägung die komplexeste wäre.3 Die nachfolgende Evolution innerhalb der Gruppe würde eine schrittweise Unterdrückung einiger "Entwicklungsschalter" bedeuten, was den Verlust der entsprechenden Module mit sich bringen würde, sowie eine zeitliche Versetzung der Ausprägung anderer Module erfordern (Bateman 1994, 568; vgl. Tab. 3, S. 569 dort). |
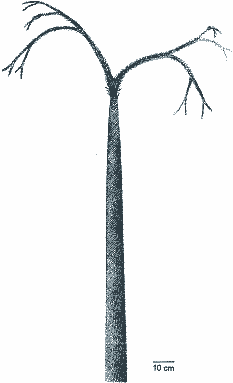
Als Ausgangsgruppe der Lepidophyten gelten die unter- und mitteldevonischen Protolepidodendrales. Dabei handelt es sich um kleine krautförmige Lycopsiden (Bärlappkräuter), denen die typischen Lepidophytenmerkmale fehlen. Übergangsformen zwischen krautigen und baumförmigen Lycopsiden sollten im Mittel- bis Oberdevon anzutreffen sein, da im Unterkarbon bereits typische Lepidophyten vorkommen. Tatsächlich sind einige Gattungen bekannt, die bezüglich mancher Merkmale einen Übergangscharakter zeigen. Schweitzer (1969) faßt im Anschluß an Hirmer (1927) solche Gattungen in der Familie Sublepidodendraceae zusammen, die zwischen den Protolepidodendrales und Lepidophyten vermitteln könnten: kleine bis mittelgroße baumförmige Lycopsiden vom Habitus der Schuppenbäume mit einfacher gebauten Blattpolstern, ohne Ligula und ohne Parichnosnarben (ein Beispiel zeigt Abb. 4). Wegen Bruchstückhaftigkeit, unterschiedlicher Erhaltungszustände und enormer intraspezifischer Variabilität der Reste ist die Familie schwer untergliederbar. Die Einteilung in Gattungen basiert auf der Blattstellung und dem Vorhandensein oder Fehlen einer echten Blattnarbe (Schweitzer 1969, 109). Die geringe Erhaltung ermöglicht nur einen "oberflächlichen Vergleich" zwischen dieser Familie und anderen Gruppen paläozoischer Lycopsiden (Jennings 1972, 80). Solange keine besser erhaltenen Formen in größerer Zahl gefunden werden, steht der Übergangscharakter dieser Familie auf schwachen Füßen. Die hauptsächlichen Merkmale der typischen karbonischen Lepidophyten (z. B. das Stigmariensystem, verschiedene Zapfentypen) treten relativ plötzlich auf. Die Lepidophyten gelten geradezu als typisches Beispiel einer saltatorischen Evolution (Thomas 1978, 323; vgl. Bateman 1994, 532).4 Thomas (1978, 323f.) hält es für denkbar, daß eine Reihe karbonischer Gattungen unabhängig (also polyphyletisch) aus dem devonischen Pool möglicher Vorläufer entstanden sind, was allerdings ein enormes Ausmaß an Konvergenzen erfordern würde. Bemerkenswert ist in diesem Zusammenhang, daß Bateman et al. (1992) für ihre cladistische Analyse (s. o.) keine geeignete Außengruppe unter den bekannten Taxa wählen konnten, sondern mit einem "hypothetischen Vorfahren" arbeiteten: Mit Hilfe von Computeranalysen rekonstruierte Bateman (1994, 565) das Aussehen eines hypothetischen Vorfahren, der als Außengruppe in Frage käme. Dieser hypothetischen Form könnte die Gattung Valmeyerodendron (Abb. 5) relativ nahe kommen. Von ihr sind jedoch die unterirdischen Organe und Fortpflanzungsorgane unbekannt, so daß eine Nähe zum hypothetischen Vorfahren der karbonischen Lepidophyten unsicher ist. Dies wird auch durch einzigartige Merkmale von Valmeyerodendron wie z. B. die Blattform in Frage gestellt. Nichtsdestotrotz bietet die Rekonstruktion eines hypothetischen Vorfahren eine zukünftige Prüfmöglichkeit anhand weiterer Funde (Bateman 1994, 565). |
Die Vielfalt der Formenkonstellationen der oberkarbonischen Bärlappbäume läßt sich leichter im Sinne eines Baukastensystems frei kombinierbarer Merkmale als im Rahmen monophyletischer Rekonstruktionen deuten. Die geschilderten Befunde sind im Rahmen des evolutionären Paradigmas unerwartet. Dessen Vorgabe verursacht eine Reihe von Fragen, die bislang im Wesentlichen nur durch Spekulationen (sprunghafte Evolution) beantwortet werden können. Bemerkenswert ist, daß hier wie auch in anderen Fällen (vgl. z. B. Shubin 1998) aufgrund unerwarteter Merkmalsverteilungen Lösungen durch ein erweitertes Verständnis morphogenetischer Prozesse während der Individualentwicklung gesucht werden. Möglicherweise können die untersuchten Lepidophyten in wenige oder nur einen Grundtyp im Sinne von Scherer (1993) zusammengefaßt werden. In diesem Falle wären die Merkmale der einzelnen Arten und Gattungen durch schnelle Radiation polyvalenter Stammformen entstanden, vielleicht ähnlich, wie Bateman & DiMichele (1994) sich das vorstellen (s.o.), die eine komplexe Ausgangsform an den Anfang stellen. Kutzelnigg (1993) beschreibt eine ähnliche Situation der Merkmalsverteilungen für die heutigen Maloideen (Kernobstgewächse) und zieht vergleichbare Schlußfolgerungen. |

|