|
|
|
|
Studium Integrale Journal |
Zu Beginn des Kambriums treten in weltweiter Verbreitung unterschiedlichste Baupläne in der Fossilüberlieferung auf, deren Vielfalt nicht befriedigend in evolutionäre Stammbäume eingeordnet werden kann (außerdem sind evolutionär zu postulierende Vorläuferformen unbekannt oder umstritten; vgl. Gould 1991). Eine ähnliche Situation stellt sich in der Überlieferung früher Landpflanzen dar, deren Vielfalt sich ebenfalls gegen Stammbaumdarstellungen sperrt. Dort lassen sich die unterschiedlichen Formen eher netzartig als baumartig verknüpfen (Junker 1996). In beiden Fällen wurde diskutiert, ob das Auftreten zahlreicher Mosaikformen durch "experimentelle Phasen" der Evolution zu deuten seien. Freie ökologische Zonen hätten einwandernden Organismen bei wenig Konkurrenz vielfältige Evolutionsrichtungen mit der Folge zahlreicher Konvergenzen erlaubt. Durch den Begriff der "experimentellen Phase" wird das Phänomen allerdings nur beschrieben, nicht aber erklärt (vgl. die Diskussion in Junker 1996, S. 74-76). Hier soll auf eine andere Schwierigkeit des Erklärungsversuchs einer "experimentellen Makroevolutionsphase" aufmerksam gemacht werden. In der Organismengeschichte gab es auch zu anderen Zeiten großräumige freie ökologische Zonen. So vermuten die Paläontologen, daß nach einem Massenaussterben an der Wende vom Perm zur Trias die Meere nahezu leer und ein "ideales Experimentierfeld für die Evolution" (Brandl 1998) waren, "vergleichbar mit den Rahmenbedingungen in den Meeren zu Beginn des Paläozoikums." Trotzdem sind keine neuen Baupläne aufgetreten wie an der Präkambrium/Kambrium-Grenze im Tierreich oder während des Devons im Pflanzenreich. Die neuerliche Entfaltung der Faunen erfolgte vielmehr auf niederem taxonomischem Niveau (Ordnungen, Familien). Vergleichbare Befunde zeigen sich auch in anderen Fällen. Offenbar sind freie Lebensräume allenfalls eine Bedingung, aber keine Erklärung für die Bildung neuer Baupläne. [Brandl R (1998) Stille Revolution in der Paläontologie. Nat. Rdsch. 51, 32-33; Gould SJ (1991) Zufall Mensch. München; Junker R (1996) Evolution früher Landpflanzen. Neuhausen-Stuttgart.] RJ |
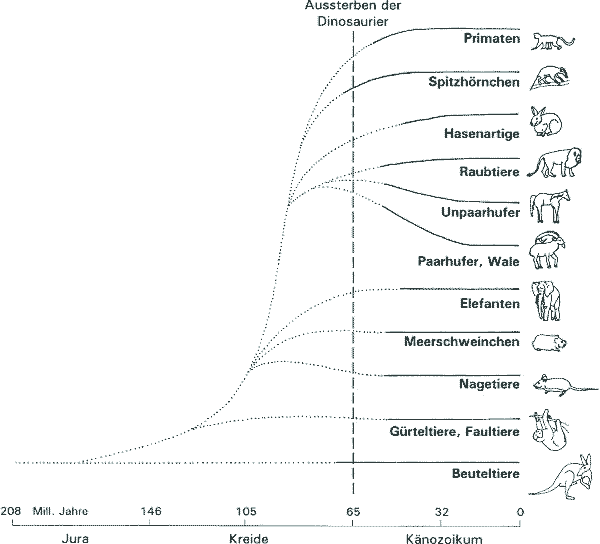
Gewöhnlich wird aufgrund von Fossilfunden davon ausgegangen, daß sich die Säugetiere erst nach dem Aussterben der Dinosaurier vor 65 Millionen Jahren in großem Umfang zu den heute bestehenden Ordnungen entwickeln konnten. Einige Spitzmaus-ähnliche Säugetiervorfahren lebten zwar bereits seit etwa 225 Millionen Jahren, sie sollen sich aber ganz im Schatten der übermächtigen Dinosaurier befunden haben. |
|
Nach einer groß angelegten Studie von Hedges & Kumar (1998) muß diese Auffassung in Frage gestellt werden. Hedges & Kumar verglichen Sequenzen von 658 Genen in 207 Tierarten. Dabei wird angenommen, daß der gemeinsame Vorfahre zweier Arten um so früher lebte, je mehr Sequenzunterschiede im Genom vorkommen. Um eine zeitliche Eichung zu ermöglichen, wurde als Fixpunkt die aus den Fossilien bekannte Trennung von säugerähnlichen und vogelähnlichen Reptilien vor 310 Jahren gewählt. Aufgrund dieser Studien zeigt sich, daß die gemeinsamen Vorfahren der meisten heutigen Säugetierordnungen (Nagetiere, Carnivoren etc.) bereits zur Zeit der Dinosaurier gelebt haben müssen. Die molekulare Uhr datiert z.B. den gemeinsamen Vorfahren aller Beuteltiere in die Zeit des Jura auf etwa vor 173 Millionen Jahre (gegenüber 95 Millionen Jahren nach der Fossilüberlieferung), und den gemeinsamen Vorfahren von Primaten und baumbewohnenden Kleinsäugern in die Kreidezeit vor etwa 86 Millionen Jahren (gegenüber 64 Millionen Jahren nach der Fossilüberlieferung), die ersten Nagetiere und Carnivoren müßten demnach vor etwa 90 Millionen gelebt haben. Das bedeutet nicht unbedingt, daß neben Tyrannosaurus und Brontosaurus Löwen und Tiger herumliefen, aber zumindest ihnen ähnliche, vielleicht körperlich kleinere direkte Vorfahren aus denselben Taxa – wofür es aber keinerlei fossile Belege gibt. Eine ähnliche Diskrepanz molekularer und fossiler Daten ist auch bei den bedecktsamigen Blütenpflanzen bekannt. Diese Studie bestätigt Ergebnisse früherer an geringeren Fallzahlen durchgeführten Untersuchungen und gilt als methodisch sehr vertrauenswürdig. Aufgrund dieser Daten muß evolutionstheoretisch eine systematische Unvollständigkeit der Fossilüberlieferung postuliert werden. [Hedges & Kumar (1998) A molecular timescale for vertebrate evolution. Nature 392, 917-920; Gibbons A (1998) Genes put mammals in age of dinosaurs. Science 280, 675-676 ] WL |
1996 wurden am Schädel eines Skelettes vom Steinzeitfriedhof Ensisheim (Elsaß) klare Hinweise auf zwei Trepanationen (chirurgisch verursachte Löcher im Schädel) entdeckt. Da die Trepanationsstellen Zeichen eines langen Heilungsprozesses aufweisen, mußten die beiden chirurgischen Eingriffe am Lebenden durchgeführt worden sein. Die technische Durchführung der Operation war meisterhaft und beweist gute anatomische Kenntnisse des Operateurs. Mit einem datierten Alter von 5.100 Jahren v. Chr. meinten Alt et al. (1997) die nachgewiesen älteste geheilte neurochirurgische Operation in der Menschheitsgeschichte veröffentlicht zu haben. Wenig später weist Lillie (1998) jedoch darauf hin, daß bereits 1966 eine noch ältere verheilte Schädeltrepanation in russischer Sprache beschrieben wurde. Der Schädel stammt aus der Region der Dnepr-Stromschnelle der Ukraine, 400 km südöstlich von Kiew. Kalibrierte Radiokarbondatierungen haben für den Fundort, den Friedhof von Vasilevka II, ein Alter von 7.300-6.220 v. Chr. bestimmt. Gegenüber dem neolithischen Ensisheimskelett stammt dieser um 1.000-2.000 Jahre ältere Schädel aus dem Mesolithikum (Mittelsteinzeit; datiertes Alter 8.000-4.000 Jahre v. Chr.). Durch die Verheilung der Trepanation ist die Methode der Knochenentfernung nicht ganz klar. Wahrscheinlich wurde aber ein Bohrer benutzt. Warum wurden in der Mittelsteinzeit Trepanationen durchgeführt? Jede Antwort darauf ist spekulativ. Am ehesten könnten die Gründe ähnlich denen heute lebender Ureinwohner Afrikas sein. Diese führen Trepanationen bei Verletzungen wie Frakturen oder Erkrankungen des Kopfes bzw. des Gehirns (andauernder Kopfschmerz, Krampfleiden, intracraniale Tumoren und Geisteskrankheit) durch. [Alt KW, Jeunesse C, Buitrago-Téllez CH, Wächter R, Boes E & Pichler SL (1997) Evidence for stone age cranial surgery. Nature 387, 360; Lillie MC (1998) Cranial surgery dates back to Mesolithic. Nature 391, 854] MB |
Kay, Cartmill & Balow (1998) haben festgestellt, daß die absolute und relative Größe des Canalis hypoglossus beim Schimpansen und Gorilla kleiner ist als beim modernen Menschen. Die Autoren vermuten einen Zusammenhang zwischen dieser Größendifferenz und der Fähigkeit zur Vokalsprache. Bei den Säugetieren verläuft durch die Schädelbasis im Canalis hypoglossalis ein Nerv (Nervus hypoglossus), der nahezu alle Muskeln der Zunge versorgt. Die menschliche Zunge sei besser mit motorischen Nerven versorgt als die Zunge der afrikanischen Großaffen. Die Ursache dafür liege in der guten motorischen Koordinationsfähigkeit der menschlichen Zunge, die Voraussetzung für die Vokalsprache ist. Von der Größe des Canalis hypoglossalis könne deshalb auf die Fähigkeit zur Vokalsprache auch bei fossilen Hominiden geschlossen werden. Der Canalis hypoglossalis liegt bei Australopithecus africanus (Sts 19, Stw 187) und möglicherweise Homo habilis (Stw 53) im Größenbereich vom Schimpansen. Beim Neandertaler (La Ferrassie, La Chapelle-aux-Saints), beim archaischen (Broken Hill, Swanscombe) und beim modernen Homo sapiens (Skuhl 5) ist er dagegen ähnlich groß wie beim heute lebenden Menschen. Kay et al. (1998) schließen aus diesem Ergebnis, daß der Mensch schon vor mindestens 400.000 Jahren eine im wesentlichen moderne menschliche Vokalsprache praktizierte. Australopithecus und möglicherweise Homo habilis besaßen dagegen wie die heute lebenden Schimpansen diese Fähigkeit nicht. Die Untersuchung von Kay et al. (1998) liefert keine Ergebnisse für Homo erectus. Dies ist sehr bedauerlich, weil Homo erectus einerseits unbestritten als echter Vertreter der Gattung Homo gilt, andererseits aber die überwiegende Zahl der Paläanthropologen ihm bis heute keine voll entwickelte menschliche Sprachfähigkeit zugestehen will. Die Möglichkeit, von der Größe eines einzigen Kanals der Schädelbasis auf das Vorhandensein einer so hochkomplexen Fähigkeit, wie sie die Vokalsprache darstellt, schließen zu können, erscheint faszinierend. Ist diese einfache Methodik aber auch stichhaltig? Kay et al. (1998) weisen selbst darauf hin, daß durch den Canalis hypoglossalis außer dem Nervus hypoglossus noch andere Strukturen (nutritive Arterien, ein Zweig der pharyngealen Arterie und ein venöser Plexus) verlaufen. Der Anteil dieser Strukturen an der unterschiedlichen Größe des Canalis hypoglossalis bei den afrikanischen Großaffen und dem Menschen bis jetzt nicht bekannt. Es ist große Vorsicht angebracht, von einzelnen anatomischen Merkmalen auf komplexe Funktionen zu schließen, wie frühere Fehlschlüsse (z.B. vom Grad der Schädelbasiskrümmung auf die Fähigkeit zur Vokalsprache beim Neandertaler) zeigen. [Kay RF, Cartmill M & Balow M (1998) The hypoglossal canal and the origin of human vocal behavior. Proc. Natl. Acad. Sci. USA 95, 5417-5419] MB |
Adolf Seilacher, Paläontologe (Tübingen / New Haven, USA) und Craförd-Preisträger (Pendant zum Nobel-Preis), beschäftigt sich seit vielen Jahren u.a. mit präkambrischen Organismen und geht neue Forschungswege. Eine alternative, umstrittene Interpretation spätpräkambrischer Lebewesen, in denen andere Forscher evolutive Ausgangsformen der Tierwelt sehen ("Ediacara-Fauna"), geht auf ihn zurück. Er deutet sie als große, vielkernige, aber einzellige, im und auf dem Sediment lebende, protoplasmagefüllte abgesteppte "luftmatratzenartige Pneus" (Vendobionta), die jedoch ein blinder Zweig der Evolution gewesen seien (Seilacher 1992; vgl. Stephan 1994). Die von ihm vor einigen Jahren initiierte Wanderausstellung "Fossile Kunst" gibt zusammen mit dem gleichnamigen Katalog (Seilacher 1995) einen allgemeinverständlichen Einblick u.a. in seine Forschungen und sucht auch ästhetischen Ansprüchen gerecht zu werden. Vielzellige, der Tierwelt zugehörige Lebewesen (Metazoen), die ein Ausgangspunkt späterer Organismen gewesen sein könnten, sieht er dagegenin den weiter ins Präkambrium (Vendian; über 1 Milliarde Jahre) zurückreichendenSandkorallen (Seilacher 1992, 1997). Als solche beschreibt Seilacher zentimeterkleine, rundliche Formen aus Sand (Protolyllia (Nemiana) simplex; Ukraine, Namibia), die wegen ihrer geringen Größe keine Einzelheiten zeigen; diese sind aber an einer größeren Art aus dem unterkambrischen Mickwitzia-Sandstein (Schweden) zu sehen (P. princeps). Differenziertere Einzelheiten zeigt eine Art aus dem Ober-Ordovizium Jordaniens (P. benderi). An morphologischen Details übertroffen werden sie durch die ebenfalls aus dem Mickwitzia-Sandstein stammenden Formen Spatangopsis costata mit "4-5 stark vorspringenden Graten, welche zur Anheftung der weichen Septen des Polypenmagensgedient haben könnten" und besonders durch S. alata, die "diese Septen sogar zu großen Propellerflügeln ausgezogen und die ektodermale Oberfläche auf ein winziges Basalkissen reduziert" hat. Von anderer Seite werden Spatangopsis-Formen aber als anorganische Bildungen bzw. als Spurenfossil betrachtet (Seilacher 1997). Es dürfte einmalig sein, daß die gleiche Metazoen-Gattung sowohl aus dem Präkambrium als auch aus Kambrium und Ordoviziumbeschrieben wird. Und: die ausdifferenziertesten Arten (der zweiten Gattung) sind nicht die jüngsten Formen. Alle diese Lebewesen sollen sich durch ein internes Skelett aus organisch verkitteten Sandkörnern passiv stabilisiert haben, das normalerweise nach dem Tod wieder zu Sand zerfiel und nur etwa infolge eines Sturms und toniger Verschüttung erhalten blieb. Seilacher betont die hypothetische Natur dieser Deutungen und die Notwendigkeit weiterer Funde, um sie zu erhärten oder zu widerlegen. Er betrachtet Sandkorallen als ausgestorbene Klasse von Hohltieren; sie verkörpern nach ihm eine früheste Form von Hartskelettbildnern, entwickelt zu einer Zeit, als Tiere noch nicht die Fähigkeit besaßen, Skelette selbst zu mineralisieren, längst vor den dazu im Ordovizium fähigen rugosen Korallen mit kalzitischem Außenskelett. [Seilacher A (1992) Vendobionta als Alternative zu den Vielzellern. Mitt. hamb. zöl. Mus. Inst. 89, 1, 9-20; Seilacher A (1995) Fossile Kunst. Albumblätter der Erdgeschichte. Korb; Seilacher A (1997) Sandkorallen – Ein ausgestorbener Lebensformtyp. Fossilien 14, 79-84; Stephan M (1994) Neuere Forschungen zur Lebewelt im Kambrium und Präkambrium – ein Überblick. Stud. Int. J. 1, 4-11] MS |
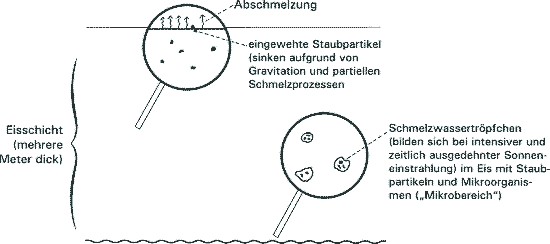
Priscu und Mitarbeiter (1998) haben in umfangreichen Untersuchungen vor allem an Eisbohrkernen aus den ca. 4 m dicken Eisdecken des Lake Bonney (Antarktis, McMurdo Valley) einen bisher wenig beachteten Lebensraum im Eis der Antarktis beschrieben. Das Eis der antarktischen Seen dieses Gebiets ist typischerweise 3-6 m dick, permanent gefroren und enthält unter der Oberfläche Schichten aus Sand und organischem Material äolischen Ursprungs. Im Sommer, während ca. 150 Tage kontinuierlicher Sonneneinstrahlung können bis zu 40% der Eisschicht schmelzen und als flüssiges Wasser vorliegen. Durch die Untersuchungen konnte ein mikrobielles Ökosystem charakterisiert werden, das aus Cyanobakterien (Phormidium, Chamaesiphon, Nostoc, nicht identifizierten coccoidalen Cyanobakterien) und Diatomeen besteht. In diesem Ökosystem dienen die Sedimentpartikel als "Mikrobereiche" mit (organischem und anorganischem) Nährstoffangebot, in denen sich ein komplexes mikrobielles Ensemble etabliert, das gleichzeitig Photosynthese betreibt, Stickstoff fixiert und organische Stoffe abbaut. Die von Priscu et al. (1998) beschriebenen Mikro-Ökosysteme unterscheiden sich von denen in marinem Packeis oder temporär mit Eis bedeckten Seen, in welchen die Mikroorganismen aus der jeweils zugrundeliegenden Wassersäule stammen (s. Literaturangaben in Priscu et al. 1998). |
|
|
Psenner & Sattler (1998) vergleichen diese Resultate in einer Diskussion mit mikrobiellen Ökosystemen im Eis von hochalpinen Seen, die in kalten Jahreszeiten eisbedeckt sind. Damit sind erneut überraschende Nischen ins Blickfeld gerückt worden, in welchen sich interessante Lebensgemeinschaften etabliert haben. Beide Autorenteams schließen die Diskussion ihrer faszinierenden Ergebnisse mit dem (künstlich und aufgesetzt wirkenden, aber sehr effektvollen) Hinweis, daß solche Befunde angesichts der Indizien für Vorkommen von gefrorenem Wasser auf Mars, dem Jupitermond Europa und anderen Himmelskörpern in Zusammenhang mit der Entstehung des Lebens gebracht werden können. Dabei ist Wasser für Lebewesen, wie wir sie kennen, eine essentielle, aber bei weitem nicht hinreichende Voraussetzung für die Existenz von (Mikro-)Organismen. Es ist zu wünschen, daß bei der Präsentation solch faszinierender Forschungsergebnisse auf Effekthascherei dieser Art verzichtet wird. Sie wird nur zu gerne von der populären Literatur aufgenommen und entpuppt sich nicht selten (vergleiche die jüngsten Publikationen zu Lebensspuren auf dem Mars) als Windei. [Psenner R & Sattler S (1998) Life at the freezing point. Science 280, 2073-2074; Priscu JC, Fritsen CH, Adams EE, Giovannoni SJ, Paerl HW, McKay CP, Doran PT, Gordon DA, Lanoil BD & Pinckney JL (1998) Perennial antarctic lake ice: an oasis for life in a polar desert. Science 280, 2095-2098.] HB |
Die Leviten waren derjenige Stamm aus den zwölf Stämmen Israels, der im allgemeinen für religiöse Angelegenheiten zuständig war. Aus diesem Stamm war es Moses Bruder Aaron, der ca. im 14. Jh. v. Chr lebte, von dem alle Priester (Kohanim) abstammen – zumindest nach den historischen Texten des Alten Testaments. Wenn diese Einteilungen sich bis heute erhalten haben, dann sollten alle modernen Nachfahren der Kohanim auf einen gemeinsamen Vorfahren zurückgehen, der auf keinen Fall nach dem Tempelbau (ca. 1000 v. Chr.) gelebt haben sollte. Dies sollte sich mit modernen Methoden der molekularen Evolutionsforschung überprüfen lassen. Nun berichtet ein Team von Anthropologen in Nature von den Ergebnissen einer solchen Überprüfung. Sie untersuchten auf dem männlich vererbten Y-Chromosom sechs Sequenzen mit hoher Mutationsrate (Mikrosatelliten) und sechs Sequenzen, deren Veränderung im untersuchten Zeitraum eher klein ist. Mit den langsam evolvierenden Sequenzen können ältere Gruppen unterschieden werden (z.B. die Kohanim von den übrigen Israeliten), während die schnell evolvierenden Mikrosatelliten sich eher zur Datierung des letzten gemeinsamen Vorfahren eignen, wenn man die Mutationsrate kennt. Diese konnte letztes Jahr mit Hilfe von tief verzweigten Familienstammbäumen abgeschätzt werden und beträgt etwa 0,2 % pro Generation für die untersuchten vier Basenpaare langen Mikrosatelliten auf dem Y-Chromosom. Und was kam dabei heraus? Die meisten Y-Chromosomen der Israeliten, die von ihrem Vater und Großvater behaupteten, daß er zu den Priestern gehöre, trugen das gleiche Muster in den langsam evolvierenden Sequenzen. Dies hebt sie von den übrigen Leviten ab, bei denen auch noch zwei andere Muster mit nennenswerter Häufigkeit vorkamen. Dies bedeutet, daß die Mehrheit der heutigen Priester-Nachfahren durchaus von einem Mann abzustammen scheinen, während die Israeliten, die sich heute als Leviten bezeichnen, wohl auch noch andere genetische Wurzeln haben können. Letzteres würde man z.B. bei Ehen von Levitinnen mit nicht-levitischen Männern erwarten, wenn deren Kinder hinterher als Leviten aufwachsen. Doch wann lebte nun der letzte gemeinsame Vorfahre aller Priester? Nimmt man obige Mutationsrate an, dann lebte Aaron etwa vor 106 Generationen. Um daraus ein absolutes Datum zu machen, müssen jedoch (i) die Länge der Generationen, (ii) die Auswirkung der Form des Stammbaums auf die Zufälligkeit der Verteilung der sich ansammelnden Mutationen und (iii) die Unsicherheiten in der Mutationsrate berücksichtigt werden. Bei Generationslängen von 25-30 Jahren ergibt sich ein Datum zwischen etwa 650 v. Chr. und 1180 v.Chr. Simulationen des Mutationsgeschehens lassen jedoch mit 95% Wahrscheinlichkeit erwarten, daß die wahre Anzahl der Generationen seit Aaron zwischen 84 und 130 liegt. Zieht man nun auch noch die Unsicherheiten in der Mutationsrate in Betracht, so könnte Aaron vor mindestens 34 und höchstens 455 Generationen gelebt haben, wobei andere Annahmen über die Form des Stammbaums zu wieder anderen Werten führen. Da beide Extremwerte (vor 850 bzw. 13650 Jahren) keinen historischen Sinn machen, bleibt zu hoffen, daß genauere Messungen der Mutationsraten und präzisere Simulationen des Evolutionsprozesses das Intervall einschränken. [Thomas MG, Skorecki K, BenAmi H, Parfitt T, Bradman N & Goldstein DB (1998) Origins of Old Testament priests. Nature 394, 138-140; Heyer E, Puymirat J, Dieltjes P, Bakker E & De Knijff P (1997) Estimating Y chromosome specific microsatellite mutation frequencies using deep rooting pedigrees. Human Molecular Genetics 6, 799-803.] LL |
Vierzig Jahre, nachdem der berühmte Archäologe Yigal Yadin die lebensgroße Statue eines Löwen im nordpalästinischen Hazor ausgegraben hat, entdeckte unlängst ein anderer Archäologe, Amnon Ben-Tor von der Hebräischen Universität eine weitere Löwenstatue. Es spricht einiges dafür, daß beide Figuren ursprünglich zusammen gehörten. Beide Löwen sind nur auf einer Seite bearbeitet, was die Nutzung als Türpfosten nahelegt. Löwenstatuen als Eingänge zu Palästen oder Tempeln waren in Syrien und Babylon, aber auch in Assyrien und im Hethiterreich weit verbreitet. Anders als im Falle des ersten Löwen, den Yadin als Türpfosten eines Tempels in der Unterstadt von Hazor identifizieren konnte, erlaubt der neue Löwe keine so eindeutige Zuordnung. Fundkontext ist ein israelitisches Gebäude aus dem 8. Jhd. v. Chr., in dessen Mauerwerk der Löwe wohl "wiederverwertet" worden ist. Das Haus stand auf dem Fundament des Königspalastes von Hazor aus kanaanitischer Zeit. Die Stadt Hazor war bis zur Landnahme durch die Israeliten so etwas wie die heimliche Hauptstadt des nördlichen Kanaan. [The pride of Hazor. Lion statue regams its long-lost mate. Biblical Archaeology Review 23 (1997), 25.] UZ |
Im dritten Buch seiner "Geschichte des judäischen Krieges" beschreibt der jüdisch-römische Geschichtsschreiber Flavius Josephus (37-100 n. Chr.) die überaus erbitterten Kämpfe, die im Jahr 67 n. Chr. nach 47tägiger Belagerung zur Einnahme der galliläischen Stadt Jodfat (auch Jotapata oder Iotape) durch die Römer geführt hatten. Der Blutzoll war gewaltig, zumal die meisten der Aufständischen lieber sterben als sich ergeben wollten. Als die Römer die Stadt schließlich durch Verrat einnahmen, konnte sich Josephus, der bis dahin die Verteidigung der Stadt geleitet hatte, mit vierzig vornehmen Einwohnern der Stadt in einer Höhle verbergen. Während die anderen Selbstmord begingen, rettete Josephus sein Leben, indem er zu den Römern überging. Obwohl die Lokalisierung des antiken Yodfat seit längeren bekannt war, wurde mit ersten Grabungen erst 1992 begonnen. Zu den Funden, die bisher von dem Team um Mordechai Aviam von der Israelischen Altertumsbehörde freigelegt wurden, gehören Hinweise auf die Schlacht selbst, wie Pfeilspitzen, daneben hasmonäische Münzen, aramäisch beschriebene Tonscherben und eine reich dekorierte Privatwohnung. Zwei mikva’ot, rituelle Bäder, weisen die Stadt eindeutig als jüdisch aus. Im Sommer 1997 entdeckte die Gruppe in einer Zisterne in unmittelbarer Nähe der Stadt Skelettreste von mindestens 30 Personen – wohl Opfer der Kämpfe vom Jahre 67. Die Skelette sind unvollständig, weshalb man vermutet, daß sie einige Jahre nach der Schlacht gesammelt und in der Zisterne beigesetzt wurden. [Rochmann B (1997) Mass grave may date to Jewish revolt. Josephus gets a Boost. Biblical Archaeology Review 23, 26.] UZ |
"SPES BONA ADIV(T)ORIB(VS) OFFICI CVSTODIAR(VM)", "Glück den Gehilfen im Wachgebäude". Diese Inschrift auf einem gut erhaltenen Mosaikfußboden im antiken Cäsarea läßt Yosef Porath von der israelischen Altertumsbehörde vermuten, daß er auf Reste des Gebäudes gestoßen ist, in dem der Apostel Paulus zwischen 58 und 60 n. Chr. gefangen gehalten wurde. Lukas schildert die Ereignisse ausführlich in der Apostelgeschichte, Kapitel 24 bis 26. Die von mehreren Grabungsteams freigelegte Ruine war früher Teil eines 15.000 m² großen Komplexes, des Praetoriums der antiken Küstenstadt. Ursprünglich von König Herodes als Residenz errichtet, umfaßte das Gelände am Meer einen großen Palast, mehrere Verwaltungsgebäude, ein Bad und weiträumig angelegte Höfe. Es grenzte an ein Amphitheater und ein Hippodrom. Caesarea war vom ersten bis zum dritten Jahrhundert n. Chr. Sitz des römischen Stadthalters in Judäa. Paulus war hier vor den römischen Statthaltern Felix und Festus und vor dem König Herodes Agrippa II. verhört worden, bevor er als Gefangener nach Rom gebracht wurde. [Rochmann B (1998) Imperial slammer identified Caesarea complex may have been Paul’s prison. Biblical Archaeology Review 24, 18.] UZ |

|