 |
Der Wurmfortsatz: Vom Nichtsnutz zum Mysterium
von Henrik Ullrich
Studium Integrale Journal
20. Jahrgang / Heft 2 - Oktober 2013
Seite 111 - 115
|
 |
Zusammenfassung: Der Wurmfortsatz, ein Anhängsel am Blinddarm, hat keinen guten Ruf. Häufig ist er Anlass für Schmerzen im rechten Unterbauch, die ein chirurgisches Eingreifen erforderlich machen. In der Evolutionsbiologie gilt der Wurmforstsatz als Paradebeispiel für ein rudimentäres Organ, welches seine ursprüngliche Form und Funktion im Verlauf der Evolution verloren hat und sich deshalb in einer zurückgebildeten Version vorfindet. Die Forschungen der letzten Jahre haben dieses traditionelle Bild vom Wurmfortsatz widerlegt. Er ist Träger wichtiger biologischer Funktionen (z. B. Infektabwehr) und zeigt eine unerwartete Verteilung unter den Säugetieren. Letztere zwingt dazu, den Wurmfortsatz aus evolutionärer Perspektive nicht mehr als eine zurückgebildete, sondern als eine mehr als 30 mal neu entstandene Struktur zu bewerten.
| |
„Wahrscheinlich haben wir immer noch nicht das Mysterium der Appendix komplett geknackt.“ So die Reaktion des Evolutionsmediziners Randolph Nesse in „Science Now“ vom 12. 2. 2013 auf die Ergebnisse einer Studie zur Verteilung des Wurmfortsatzes (Appendix) unter Säugetieren und dessen Evolution. Dieses Anhängsel an unserem Blinddarm (Caecum) hat bekanntlich keinen guten Ruf. Denn er wird fast ausschließlich mit Schmerzen im rechten Unterbauch und mit chirurgischem Eingreifen in Verbindung gebracht. Als zweckloser Überrest unserer Stammesgeschichte steht er in fast allen Evolutions-Lehrbüchern. Nun soll er ein „Mysterium“ sein? Wie kam es zu diesem erstaunlichen Wandel bezüglich der Beurteilung des Wurmfortsatzes, dem kleinen Anhängsel unseres Dickdarms (Abb. 1)?
|
Unvollkommen, zwecklos und rudimentär
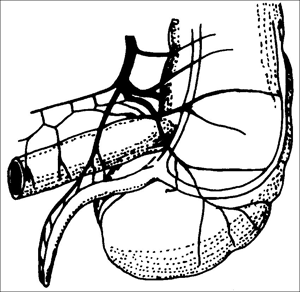 |
| Abb. 1: Schemazeichnung des menschlichen Wurmfortsatzes. Schon die gute Durchblutung deutet auf Funktionalität hin. (Nach Murris 1991) |
|
Charles Darwin befasste sich in seinem Hauptwerk „Die Entstehung der Arten durch natürliche Zuchtwahl“ in einem Abschnitt über „Rudimentäre, atrophierte und abortive Organe“. Darin erläutert er: „Organe oder Theile, in diesem eigenthümlichen Zustande den Stempel der Nutzlosigkeit tragend, sind in der Natur äusserst gewöhnlich. ... Ich glaube, dass Nichtgebrauch dabei hauptsächlich in Betracht komme, der während einer langen Generationen-Reihe die allmähliche Abschwächung der Organe veranlassen kann, bis sie endlich nur noch als Stümmel erscheinen. ...
Nach der Annahme einer Fortpflanzung mit Abänderung können wir schliessen, dass das Vorkommen von Organen in einem verkümmerten, unvollkommenen und nutzlosen Zustande und deren gänzliches Fehlschlagen, statt wie bei der gewöhnlichen Theorie der Schöpfung grosse Schwierigkeiten zu bereiten, vielmehr vorauszusehen war und aus den Erblichkeits-Gesetzen zu erklären ist“ (Übersetzung der 2. Auflage nach H. G. Bronn, 1860, S. 453, 458, 460).
So schrieb Charles Darwin dem Wurmfortsatz des Menschen und zugleich auch den wenigen bis dahin bekannten analogen Strukturen bei einigen Säugetieren und Großaffen keine biologische Funktion mehr zu. Nach seiner Hypothese führte ein verändertes Nahrungsverhalten unserer Vorfahren dazu, dass sich der einst größere Blinddarm deutlich verkürzte. Der Blinddarm ist der erste Teilabschnitt des Dickdarms. Darwin nahm weiter an, dass der Wurmfortsatz (Appendix) offensichtlich eine geschrumpfte, zurückgebildete Tasche (Divertikel) des früheren langen Blindarms repräsentiert. Zur Zeit Darwins existierten nur wenige Daten zum Vorkommen des Wurmfortsatzes bei den Säugetieren. Seine Schlussfolgerungen waren unter Zugrundelegung seiner Abstammungslehre und Vererbungslehre nachvollziehbar und eine willkommene, eingängige Bestätigung seiner Thesen.
Als Paradebeispiel für ein rudimentäres bzw. stammesgeschichtlich rückgebildetes Organ hielt diese Deutung des Wurmfortsatzes fast unverändert unter Berufung auf Darwins Hypothese Einzug in viele Publikationen und Lehrbücher als Beleg der menschlichen Stammesgeschichte. So kann man z. B. aktuell in Meyers Großes Konversations-Lexikon dazu lesen (Stand 8. 7. 2013):
„Rudimentäre Organe des Menschen, Körperteile, die ihres verkümmerten Zustandes wegen fast oder vollkommen leistungsunfähig sind. Sie sind aufzufassen als letzte Überreste von Organen, die bei den Vorfahren des Menschen wohl entwickelt und leistungsfähig waren, allmählich aber infolge von Nichtgebrauch im Laufe der Generationen rückgebildet wurden. …
Schließlich [sei der] … sogen. W u r m f o r t s a t z des Darmes erwähnt, der den verkümmerten Rest eines ehemals stärker entwickelten Blinddarmes darstellt und jetzt nur noch als Sitz gefährlicher Erkrankungen von Bedeutung ist“ (http://tinyurl.com/kq22ef9).
|
In seiner späteren Abhandlung „Die Abstammung des Menschen“ (3. Auflage, 1875) nutzte Darwin die evolutionäre Herangehensweise, um sein „Argument der Unvollkommenheit“ am Beispiel des Wurmfortsatzes zu erhärten: „In Bezug auf den Verdauungskanal ist mir nur ein einziges Beispiel von einem Rudimente vorgekommen, nämlich der wurmförmige Anhang des Blinddarms. Der Blinddarm ist eine Abzweigung oder ein Divertikel des Darms, welcher mit einem Blindsack endigt, und bei vielen niedrigeren pflanzenfressenden Säugethieren ist er ausserordentlich lang, bei dem marsupialen Koala ist er factisch über dreimal so lang als der ganze Körper. Zuweilen ist er in einen langen, sich allmählich zuspitzenden Fortsatz ausgezogen und zuweilen in Abtheilungen abgeschnürt. Es scheint, als wenn in Folge veränderter Ernährung oder Lebensweise der Blindsack bei verschiedenen Thieren sehr verkürzt worden sei, wo dann der wurmförmige Anhang als Rudiment des verkürzten Theils übrig bleibt. Dass dieser Anhang ein Rudiment ist, können wir aus seiner unbedeutenden Grösse und aus den Beweisen für seine Veränderlichkeit beim Menschen schliessen, welche Professor Canestrini gesammelt hat. Er fehlt gelegentlich vollständig oder ist wiederum bedeutend entwickelt; seine Höhle ist zuweilen vollständig für die Hälfte oder zwei Drittel seiner Länge verschlossen, wobei dann der Endtheil aus einer abgeplatteten, soliden Ausbreitung besteht. Beim Orang ist dieser Anhang lang und gewunden; beim Menschen entspringt er vom Ende des kurzen Blinddarms und ist gewöhnlich 4-5 Zoll lang, während er nur ein Drittel Zoll im Durchmesser hat. Er ist nicht bloss nutzlos, sondern wird zuweilen Todesursache, von welcher Thatsache mir vor Kurzem zwei Fälle bekannt geworden sind. Es rührt dies daher, dass kleine, harte Körper in den Kanal eindringen und dadurch Entzündung verursachen“ (Übersetzung der 3. Auflage nach C. G. Carus, 1875 Band I, S. 26).
|
|
|
|
Der Wurmfortsatz: Zufluchtsort und Schutzschild
Die Forschungen insbesondere der letzten Jahrzehnte haben diese Beurteilungen jedoch vollständig widerlegt. Das gilt für die behauptete Unvollkommenheit, die vermutete Funktionslosigkeit, das Auftreten des Wurmfortsatzes unter den Säugetieren und damit auch für dessen evolutionäre Geschichte. Wenn man letztere zugrundelegt, muss sie komplett umgeschrieben werden.
Seit mehr als einem Jahrhundert ist bekannt, dass der feingewebliche Aufbau des Wurmfortsatzes reich an Lymphgewebe ist (z. B. Berry 1900). Die darin sich befindlichen weißen Blutzellen besitzen eine zentrale Funktion bei der Abwehr von Infektionen. Nicht selten bezeichnet man deshalb den Wurmfortsatz auch als Darmmandel (bzw. Darmtonsille). In den letzten Jahren zeigte sich zusätzlich eine enge funktionelle Verknüpfung zwischen dem menschlichen Wurmfortsatz und der Besiedlung des Dickdarms mit lebenswichtigen Darmbakterien. Diesen Bakterien fällt die Aufgabe zu, die Ausbreitung gefährlicher Krankheitserreger im menschlichen Verdauungstrakt zu verhindern. Forscher fanden heraus, dass die Appendix des Menschen eine Art Zufluchtsort („safe house“) und Rettungsstation für symbiotische Bakterien darstellt (Bollinger 2007), die das Wachstum nützlicher Darmbakterien fördert und bei durchfallbedingten Darmentleerungen die Wiederbesiedlung mit diesen Bakterien ermöglicht bzw. erleichtert (vgl. dazu auch Junker 2003, 2008). Ein dünner Biofilm kleidet das innere Lumen (Hohlraum) des Wurmforstsatzes aus und bildet den natürlichen Nährboden für diese Darmbakterien. Bei der Aufarbeitung von Daten von Patienten nach Entfernung des Blinddarms zeigte sich auch ein enger physiologischer Zusammenhang hinsichtlich der Entstehung von chronisch entzündlichen Darmerkrankungen (wie z. B. Colitis ulcerosa, Morbus Chron) und dem Vorhandensein des Wurmfortsatzes (Smith et al. 2009, 2013).
Dieses Wissen und das im Weiteren noch zu erläuternde relativ häufige Auftreten von Wurmfortsätzen bei den Säugetieren macht zweifelsfrei klar, dass diesem kleinen Organ eine wichtige physiologische Bedeutung zukommen muss und es für die bisher beschriebenen Funktionen optimal konstruiert ist (Laurin et al. 2011).
„Die Datenlage stützt die Einsicht, dass die Appendix des Caecums eine immunologisch relevante Struktur ist. Ihre primäre Funktion ist die eines Zufluchtsortes für nützliche Bakterien. Dies basiert auf den Ergebnissen verschiedener Disziplinen einschließlich der Medizin, der Epidemiologie, Immunologie und Mikrobiologie ...“ (Smith 2013, 15).
|
|
Rudiment der Stammesgeschichte?
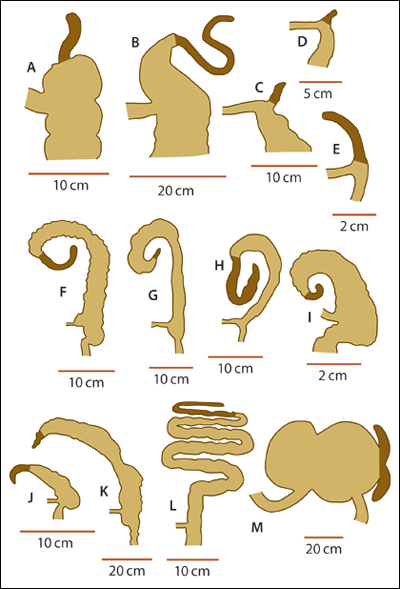 |
| Abb. 2: Morphologie der Appendix bei verschiedenen Säugetieren. Die Appendix ist jeweils dunkelbraun gezeichnet. A Mensch, B Orang-Utan, C Wombat, D Ameisenigel, E Schnabeltier, F Kaninchen, G Bodencuscus, H Opossum, I Kap-Standgräber (Graumull), J Stachelschwein, K Biber, L Koala, M Seekuh. (Nach Smith 2013 |
|
Wenn dem Wurmfortsatz eine wichtige funktionelle Bedeutung zukommt, ist dann seine phylogenetische Deutung als rudimentäres Organ noch aufrecht zu erhalten? Charles Darwin wies im oben genannten Kapitel seines Hauptwerkes „Über die Entstehung der Arten“ auch darauf hin, dass rudimentäre Organe durchaus Restfunktionen oder auch in einem morphologisch zurückgebildeten Stadium neue Funktionen übernehmen können. In dieser eingeschränkten Weise wäre also dessen Wertung als Rudiment in Anlehnung an Darwin noch möglich.
Die systematische Verteilung des Auftretens des Wurmforstsatzes bei den Säugetieren, das nachweisbare Fehlen einer Abhängigkeit seiner Existenz von Ernährungsgewohnheiten des jeweiligen Trägers und das Fehlen einer Korrelation von Ernährung und Länge des Blinddarms widersprechen jedoch deutlich dieser Deutung. Im Folgenden werden diese drei Argumente erläutert.
Rebecca E. Fisher von der Yale School of Medicine wies bereits 2000 in einer Arbeit darauf hin, dass die klassische stammesgeschichtliche Bewertung des Organs sich nicht mit den anatomischen und systematischen Fakten deckt. Die Auffassung, es handle sich bei der Appendix um ein rückgebildetes Organ, beruht ihrer Auffassung nach auf dem Vergleich mit solchen Säugetieren, die einen großen Blinddarm (z. B. Koala, Hauspferd) oder eine lange Appendix besitzen (z. B. Hase). Zusätzlich fehlten bislang, so Fisher weiter, klare Kriterien, die den Wurmforstsatz als solchen definieren. Als Kriterien für die Definition der Appendix schlug sie deshalb vor (vgl. Abb. 2):
• Eine schmale, markante Spitze (Apex), welche dem übrigen Blinddarm an seinem vorderen Ende aufsitzt,
• ein deutlich dickwandiger Aufbau dieser Spitze (Apex) im Gegensatz zum dünnwandigen Blinddarm und
• Konzentration von lymphoidem Gewebe (Gewebestrukturen, die reich sind an Lymphozyten, einer Untergruppe der weißen Blutkörperchen).
Auf der Grundlage dieser standardisierten morphologischen Kriterien ergab sich ein völlig anderer Blick auf das Vorkommen der Appendix innerhalb des Systems der Säugetiere (Fisher 2000, 229). Die daraus abzuleitenden phylogenetischen Interpretationen änderten sich dramatisch. Denn die Wurmfortsätze der höheren Primaten erschienen abgeleiteter („höherentwickelter“) als die der niederen Primaten – die Verhältnisse sind also genau umgekehrt wie nach evolutionstheoretischen Erwartungen. Allerdings waren die Verhältnisse bei vielen Primaten noch ungenügend bekannt. Vor einer stabileren Beurteilung forderte Fisher daher weitere Untersuchungen.
2007 sah Bollinger trotz der bis dahin immer noch vermuteten geringen Verbreitung des Wurmfortsatzes die These von Fisher bestätigt: „Das sporadische Vorkommen der Appendix in der Stammesgeschichte dürfte nahelegen, dass diese Struktur eher evolutionär abgeleitet ist und einer spezifischen Funktion dient, als in ihr ein bloßes Überbleibsel eines vormals wichtigen Verdauungsorgans zu sehen“ (Bollinger et al. 2007, 827).
Neue Ergebnisse, die zu einer völlig neuen Bewertung der Evolution des Wurmfortsatzes führten, lieferten die Studien von Smith et. al. (2009, 2013). Die gezielte Analyse der Anatomie zahlreicher heute lebender Säugetiere offenbarte bei unerwartet vielen Arten einen Wurmfortsatz. In den Studien von Smith et al. wurden die von Fisher formulierten Erkenntnisse bestätigt. Darüber hinaus forderten die Autoren hinsichtlich der evolutionären Geschichte des Wurmfortsatzes eine komplett neue Sicht ein. Smith et al. gingen 2009 noch davon aus, dass der Wurmfortsatz mindestens zweimal unabhängig (konvergent) entstanden sei, einmal in der Gruppe der Marsupialia (Beuteltiere) und in den Euarchontoglires, einer Untergruppe der Säugetiere, zu denen u. a. die Primaten, Hasenartigen und die Spitzhörnchen gehören.
Bei ihrer aktuellen Analyse (2013) von 361 lebenden Säugetieren fanden sich 50 Arten, die über einen Wurmfortsatz verfügen. Neben dem Schnabeltier (Monotremata) und einigen Beuteltierarten (z. B. Opossum und Wombat) besitzen z. B. Kaninchen, Biber, Koalabären und das Stachelschwein eine Appendix exakt an der gleichen Position wie beim Menschen mit analogem feingeweblichen Aufbau, jedoch mit großer anatomischer Variabilität (vgl. Abb. 2). Das heißt, der Wurmfortsatz ist unter den Säugetieren deutlich weiter verbreitet, als man noch bislang angenommen hatte. Fügt man die nun bekannte Verteilung seines Auftretens in den aktuellen, auf molekularbiologischen Daten beruhenden Stammbaum der Säugetiere (Bininda-Emonds et al. 2007) ein, ergibt sich daraus eine unerwartete Schlussfolgerung: Es muss in mindestens 32 Fällen von einer konvergenten, also unabhängigen Entstehung des Wurmfortsatzes bei den Säugetieren ausgegangen werden. (vgl. Abb. 3). Zusätzlich – so eine weitere Folgerung aus der phylogenetischen Interpretation dieser Merkmalsverteilung unter den Säugetieren – scheint der Wurmfortsatz bei maximal 7 Tierarten evolutionär zurückgebildet zu sein.
Damit ist entgegen aller Erwartungen gezeigt, dass der Wurmfortsatz ein abgeleitetes (apomorphes, neues) und kein rudimentäres Merkmal ist. Der zweite ebenso wichtige Schluss ist der, dass trotz der großen Ähnlichkeit der Wurmfortsatz innerhalb der Säugetiergruppen kein homologes, sondern ein konvergentes Merkmal repräsentiert. Bei den bisher diesbezüglich 361 untersuchten Säugetieren ergibt sich eine mehr als 30-fach unabhängige Entstehung (s. o.). Es ist damit zu rechnen, dass bei der Untersuchung weiterer der heute bekannten ca. 5500 Säugetierarten diese Zahl noch steigen wird.
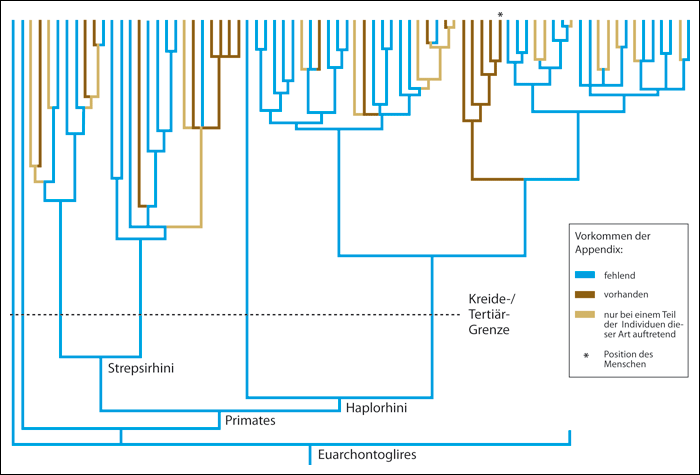 |
| Abb. 3: Cladogramm der Euarchontoglires. Bei dieser Gruppe (für die es keine deutsche Bezeichnung gibt) handelt es sich um eine Überordnung der höheren Säugetiere, die die Nagetiere, Hasenartigen, Spitzhörnchen, Primaten und Riesengleiter umfasst. Allein in dieser Gruppe muss vielfach unabhängige Entstehung der Appendix angenommen werden (braune Äste). Jeder Ast steht für eine Art aus dieser Gruppe. (Nach Smith et al. 2013; Details siehe dort) |
|
|
|
Ernährung und Länge des Blinddarms ohne Einfluss
Einer zweiten Frage gingen Smith et al. in ihrer aktuellen Studie nach. Gibt es tatsächlich einen Zusammenhang zwischen den Ernährungsgewohnheiten und dem Auftreten des Wurmfortsatzes in den einzelnen Säugetierlinien, wie nach Darwins Selektionstheorie erwartet wurde?
Auch dieser Deutungsansatz wurde widerlegt. Es fand sich bei der Betrachtung aller untersuchten Säugetiere kein Hinweis dafür, dass die Evolution des Wurmfortsatzes als eine Reaktion auf Veränderungen von Ernährungsgewohnheiten oder des sozialen Verhaltens erklärbar wäre. Die bei Primaten zwar richtige Beobachtung Darwins, dass ein vergleichsweise kurzer Blinddarm mit der Existenz des Wurmfortsatzes korreliert, lässt sich nicht auf die Gesamtheit der Säugetiere übertragen. Die Autoren sind deswegen der Ansicht, dass in jeder einzelnen Entwicklungslinie andere multifaktorielle Ursachengefüge dessen Entstehung oder sein Verschwinden hervorriefen.
|
|
Mysterium Appendix: Die Zeit ist reif für eine neue Hypothese
In ihrer abschließenden Diskussion kommen Smith et al. zu einer interessanten Schlussfolgerung: „Es erscheint einsichtig, dass wenn Darwin Zugriff auf mehr Fakten bezüglich des Auftretens der Appendix am Caecum bei Säugetieren gehabt hätte, er nicht das evolutionäre Szenario der Appendix so beschrieben hätte wie in seinem bahnbrechenden Werk. Wenn man sich das klar macht, scheint die Zeit reif dafür zu sein, eine neue Hypothese über die Entstehung der Appendix am Caecum zu formulieren“ (Smith et al. 2013, 14).
 |
| Der Wurmfortsatz ist ein Beispiel dafür, dass die biologische Forschung nicht zu den evolutions-theoretisch prognostizierten Ergebnissen führte. |
 |
|
Der Wurmfortsatz ist ein weiteres Beispiel dafür, dass die durch Darwin motivierte biologische Forschung nicht zu den evolutionstheoretisch prognostizierten Ergebnissen führte. Das „Argument der Unvollkommenheit“ oder der evolutionär bedingten Konstruktionsfehler verliert mit der Klärung der funktionellen Bedeutung des Wurmfortsatzes einen weiteren wichtigen Kronzeugen (vgl. zum Argument des Konstruktionsfehlers auch Ullrich in dieser Ausgabe).
Die bisher zu vermutende 30-fache unabhängige Entstehung der Appendix als neues Organ in unterschiedlichen Säugetierlinien reiht sich ein in die vielen Merkmale, die im Verlauf der Evolution bei Säugetieren ebenfalls konvergent entstanden sein sollen (z. B. Merkmale der Netzhaut, Mittelohr, Flugfähigkeit u. v. a. m., Ullrich 2010, 2011, 2012). Keines der bisher diskutierten evolutionstheoretischen Modelle ist bislang in der Lage, eine kausale Erklärung für diese immer zahlreicher werdenden Fälle von Konvergenzen zu liefern.
Darwins Vererbungstheorie, die auf der Weitergabe erworbener Merkmale beruhte, war schon Anfang des 20. Jahrhunderts widerlegt, die Selektionstheorie als Motor und Dirigent der Evolution konnte ihre Ansprüche nur in einem engen Rahmen einlösen.
Die neuen Evo-Devo-Ansätze gewinnen ihre Attraktivität aus dem Versagen der bisherigen Theorien und dem immensen Wissenszuwachs der experimentellen Embryologie und Molekularbiologie, ohne jedoch selbst die bestehenden Fragestellungen bisher grundlegend beantworten zu können. Die Zeit ist reif, nicht nur über eine neue Hypothese der Evolution des Wurmfortsatzes nachzudenken. Die hier vorgestellten und viele andere biologische Fakten geben Anlass, alternative Modelle zur Geschichte des Lebens ernst zu nehmen. Andere Antworten wie Schöpfung sind möglich und plausibel. Auch wenn man dabei nicht nur auf dem Boden der Naturwissenschaft nach Antworten für das Mysterium des Wurmfortsatzes suchen muss.
|
Literatur
- Boisvert CA, Joss JMP & Ahlberg PE (2013)
- Comparative pelvic development of the axolotl (Ambystoma mexicanum) and the Australian lungfish (Neoceratodus forsteri): conservation and innovation across the fish-tetrapod transition. EvoDevo 2013, 4:3, doi:10.1186/2041-9139-4-3
- Freitas R, Gómez-Marín C, Wilson JM, Casares F & Gómez-Skarmeta JL (2012)
- Hoxd13 Contribution to the Evolution of Vertebrate Appendages. Dev. Cell, dx.doi.org/10.1016/j.devcel.2012.10.015
- Schneider I & Shubin NH (2012)
- Making Limbs from Fins. Dev. Cell, dx.doi.org/10.1016/j.devcel.2012.11.011
|
|  |
