 |
|
von Reinhard Junker
Studium Integrale Journal
12. Jahrgang / Heft 1 - Mai 2005
Seite 11 - 18
|
 |
Zusammenfassung: Der Übergang von den devonischen zu den karbonischen Tetrapoden ist durch eine markante Fossillücke („Romers Lücke“) fossil fast nicht bekannt. Im Karbon tauchen unterschiedlichste Mosaikformen plötzlich in großer Fülle und so großer Verschiedenartigkeit auf, daß Stammbaumrekonstruktionen kaum möglich sind. Die Selektionsdrücke, die zur Entstehung der Vierbeinigkeit bei den devonischen Formen geführt haben könnten, sind unklar. Es gibt einander widersprechende Argumente sowohl für eine Entstehung von Fingern und Tetrapodenextremität im Wasser als auch für deren Entstehung auf Land, was ein sekundäres Wasserleben mancher Tetrapoden bedeuten würde. Die jeweiligen Argumente stellen beide Szenarien deutlich in Frage, woraus ein evolutionstheoretisches Dilemma folgt. Es wird auch diskutiert, daß die oberdevonischen/ unterkarbonischen Formenabfolgen eine ökologische Zonierung anstelle eines evolutionstheoretischen Szenarios widerspiegeln könnten.
| |
|
In den beiden vorangegangenen Folgen des Artikels wurden die oberdevonischen Tetrapoden (Vierbeiner) vorgestellt. Es wurden Argumente dafür genannt, daß diese Tetrapoden den Sprung ans Land nicht vollzogen haben. Denn alle devonischen Tetrapoden weisen deutliche Einrichtungen für ein hauptsächliches oder dauerhaftes Leben im Wasser auf. Das gilt selbst für Ichthyostega, das berühmte „erste Amphib“, das mit zunehmender Kenntnis als eher wasserlebend eingestuft wurde. Im folgenden wird ein kurzer Überblick über die auf das Devon folgende Fossildokumentation im Unterkarbon gegeben. Anschließend werden einige Fragen zu Selektionsdrücken und andere evolutionstheoretische Probleme diskutiert.
|
|
Der Sprung zu den karbonischen Tetrapoden
Nach dem Oberdevon folgt eine markante Überlieferungslücke von ca. 20 Millionen Jahren in der Fossildokumentation der Tetrapoden, die nach dem berühmten Wirbeltierpaläontologen Alfred S. Romer als „Romer-Lücke“ bezeichnet wird (vgl. Abb. 20). Erst vor kurzem wurde die Gattung Pederpes als erstes Fossil beschrieben, das in diese Lücke fällt (Clack 2002b). Von der Merkmalskombination paßt dieses Fossil nur teilweise in evolutionstheoretische Vorstellungen. Im Viséum, der zweiten Stufe des Karbons, erscheint fossil dann eine Fülle sehr verschiedener Tetrapoden (Abb. 20), darunter auch eindeutig landlebende. Das Spektrum reicht von zarten, mausgroßen Tieren wie Casineria bis zu großen, räuberisch lebenden Formen wie Crassigyrinus (Abb. 22) oder beinlosen, schlangenförmigen Tieren wie die Aïstopoda (Abb. 21). Der „Konservativismus“ der oberdevonischen Formen ist aufgebrochen (Clack 2002a, 209).
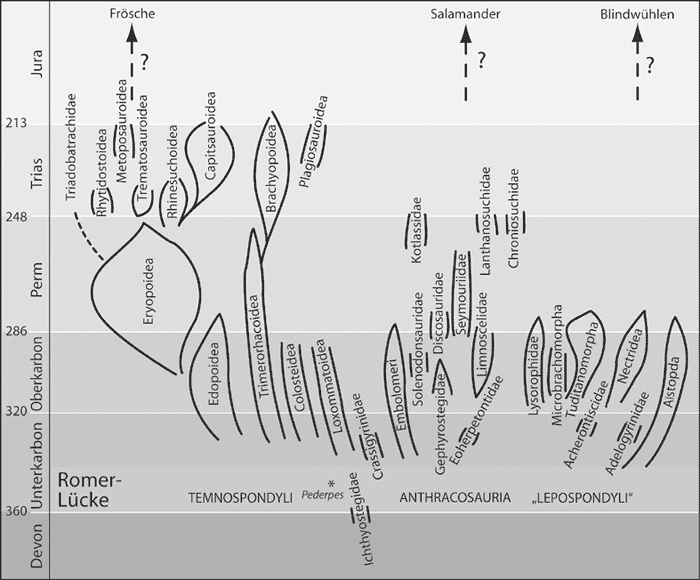 |
| Abb. 20: Überblick über die stratigraphischen Positionen der paläozoischen Tetrapoden. Die Romer-Lücke ist hervorgehoben. Die Verwandtschaftsverhältnisse können kaum bestimmt werden. Es gibt für keine Gruppe einen Konsens über die genauen Verwandtschaftsbeziehungen. Zudem sind unter den ältesten karbonischen Fossilien auch ausgesprochen abgeleitete Formen wie die Aïstopoda (Abb. 21). Schließlich sind auch die Verbindungen zu den heutigen Amphibiengruppen unklar. (Nach Carroll 1993) |
|
Manche Formen wie die Aïstopoda (Abb. 21) oder Nectridea sind so sehr spezialisiert, daß evolutionär eine lange Vorgeschichte angenommen werden muß, für die es jedoch keine Fossildokumentation gibt (Clack 2002a, 221). Nach dem Fossilbefund tauchen landlebende Tetrapoden also zugleich abrupt und in großer Verschiedenartigkeit auf. Die frühen karbonischen Tetrapoden weisen eine Diversität von Formen und Funktionen und entsprechende ökologische Verhältnisse und Lebensweisen auf, die heute nicht mehr anzutreffen ist. In vielen Fällen können über ihre evolutionäre Rolle nur Mutmaßungen angestellt werden. Ihre Verwandtschaftsverhältnisse zueinander oder zu jüngeren Formen und auch die Frage, ob sie von mehr ans Landleben angepaßten Vorfahren abstammen, sind ungeklärt (Clack 2002a, 196, 221). Viele Merkmale weisen Konflikte (Inkongruenzen) in ihrer Verteilung auf (Clack 2002a, 14).1
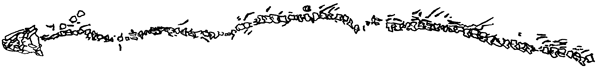 |
| Abb. 21: Die extrem spezialisierte Aïstopoden-Gattung Lethiscus; der älteste Aïstopode aus dem Viséum (mittleres Unterkarbon). (Nach Carroll 1992) |
|
Vor diesem Hintergrund ist verständlich, daß die Formenvielfalt, die ab dem Viséum im Karbon plötzlich auftaucht, kaum eine phylogenetische Rekonstruktion erlaubt (Carroll 1992, 47; Daeschler 2000, 301). Cladistische Analysen decken eine ungewöhnlich große Zahl an Konvergenzen (Mehrfachentstehung baugleicher Merkmale) und Reversionen (Rückentwicklungen) auf. Die Fülle der karbonischen Amphibiengruppen kann auf der Basis abgeleiteter Merkmale nicht in Form ineinandergeschachtelter Gabelungen phylogenetisch angeordnet werden, ohne daß eine große Anzahl von Rückentwicklungen angenommen wird (Carroll 1982, 102f.).2
Abgeleitete Formen am Anfang. Die Romersche Lücke erscheint umso markanter, als die ersten Formen, die am oberen Ende dieser Lücke fossil erscheinen, abgeleitete Merkmale aufweisen. Dies gilt insbesondere für die Aïstopoda, schlangenförmige, extrem spezialisierte Organismen (Abb. 21). Es ist nicht übertrieben zu sagen, daß man solche Formen zu allerletzt am „karbonischen Ufer“ der Romer-Lücke erwartet hätte. Aber auch sonst sind die unterkarbonischen Formen aus dem Visé und unteren Namur relativ fortschrittlich; sie ähneln späteren karbonischen Formen in ihrer taxonomischen Zusammensetzung (Ahlberg & Milner 1994, 513; vgl. Milner & Sequeira 1994, 359). „Thus, more advanced tetrapod morphologies are being described from the Viséan, while Famennian tetrapods prove to be more primitive than previously suggested“ (Ahlberg & Milner 1994, 513). Das bedeutet, daß durch die neuere Forschung die Romer-Lücke von beiden Seiten her eher tiefer geworden ist. Die einzige Ausnahme bildet Pederpes (s. o.).
Ebenfalls bereits im Viséum wurden Gattungen gefunden, die zu den beiden Hauptgruppen der Tetrapoden, den Amphibien und den Amnioten, gestellt werden. Deren evolutionärer Ursprung muß entsprechend früher angesetzt werden und ist fossil nicht dokumentiert (Clack 1994, 375f.).
|
|
Evolutionstheoretische Probleme
Sind einige oberdevonische Tetrapoden Rückkehrer ins Wasser?
In der vorausgegangenen Folge dieser Artikelserien wurde bereits erwähnt, daß Acanthostega und sogar die stratigraphisch frühe Gattung Elginerpeton, eventuell auch andere Formen, von manchen Forschern als möglicherweise sekundär wasserlebend betrachtet werden. Das heißt: Sie seien von einer früheren Landlebensweise wieder ins Wasser zurückgekehrt (Clack 2002a, 129, 135; Westenberg 1999).
Argumente für sekundäres Wasserleben.
Als Argumente für eine sekundär wasserlebende Lebensweise werden genannt:
- Die Ausbildung eines tetrapodenartigen Beckens und großer Oberschenkelknochen sowie der Besitz von Fingern. Es ist unverstanden, weshalb im Wasser diese Strukturen entstehen sollten (Clack 2002a, 135).
- Der Verlust von Stützelementen der Flossen. Warum sollten diese im Wasser verloren gehen?
- Acanthostega hat eine Abfolge von Knochen am hinteren Ende des Schädels verloren (Operculogular-Serie). Im Wasser sind diese Knochen für Kiemenatmung aber wichtig, zumindest nützlich. Die heutigen luftschnappenden Lungenfische haben trotz Besitz von Kiemen diese Knochenserie ebenfalls nicht, aber sie atmen zusätzlich mit Lungen. Auch bestimmte moderne Knochenfische (Teleostei) besitzen keine Operculogular-Knochenserie, einige von ihnen atmen Luft, andere dagegen nicht (Clack 2002a, 135f.).
- Für ein sekundäres Wasserleben einiger Tetrapoden sprechen auch physiologische Gründe. Janis & Farmer (1999, 117ff.) halten es für unwahrscheinlich, daß im Wasser Kiemen zurückgebildet oder gar aufgegeben werden, weil sie nicht nur der Atmung, sondern auch der Ausscheidung von Kohlendioxid und Ammoniak sowie der Ionenregulation und Säure-Basen-Regulation dienen. Die Ausscheidung ist eine fundamental wichtige Fähigkeit der Kiemen wasserlebender Organismen, auch wenn zusätzlich Ausscheidung über die Nieren erfolgt. „Gills are simply too valuable to be lost by an aquatic animal, even in those air-breathing fishes that no longer use the gills for oxygen uptake“ (Janis & Farmer 1999, 123). Wasserlebende Tetrapoden ohne Kiemen müßten daher nach Auffassung dieser Autoren von Formen abstammen, die zuvor bereits an Land lebten.
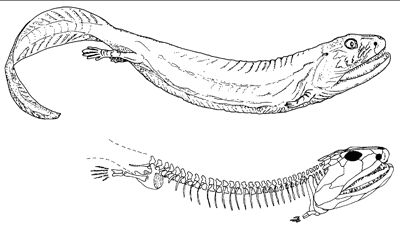 |
| Abb. 22: Crassigyrinus aus dem Viséum (mittleres Unterkarbon); vermutlich sekundär wasserlebend. (Nach Clack 2002a) |
|
Bei Acanthostega und Ichthyostega ist der Besitz von Kiemen zwar praktisch sicher, jedoch sprechen die erwähnten anatomischen Merkmale im hinteren Kopfbereich für eine Reduktion der Kiemen. Nach der Argumentation von Janis & Farmer (1999) ist aber bereits eine Rückbildung der Kiemen bei dauerhaft wasserlebenden Formen mindestens nicht naheliegend.
Argumente für primäres Wasserleben.
Einige anatomische Merkmale sprechen dagegen deutlich für eine primär aquatische Lebensweise (man müßte sonst gravierende Rückentwicklungen annehmen):
- Die Anatomie der Arme. Die Arme von Acanthostega sind Eusthenopteron am ähnlichsten, wären also am leichtesten direkt von einem Fischvorfahren abzuleiten. (Allerdings gilt die Ausbildung von Fingern als evolutionäre Neuheit, die übergangslos auftritt.) Beiden gemeinsam ist eine deutlich unterschiedliche Länge von Speiche und Elle. Bei landlebenden Formen sind diese beiden Knochen etwa gleich lang. Dagegen hat kein sekundär wasserlebender Tetrapode eine Speiche, die deutlich länger als die Elle wäre, wie dies bei Acanthostega der Fall ist (Clack 2002a, 136).
- Die Anatomie des fischartigen Schwanzes mit langen Flossenstrahlen, die an Land wenig Sinn machen (Clack 2002a, 136).
- Wenn manche wasserlebende Tetrapoden von überwiegend landlebenden abstammen würden, wäre zu erwarten, daß die Gelenke beweglicher sind als dies tatsächlich bei den oberdevonischen Tetrapoden der Fall ist (soweit die Extremitäten fossil überliefert sind).
Neben diesen anatomischen Gründen gibt es aus evolutionstheoretischer Sicht noch indirekte Argumente gegen die sekundär wasserbewohnende Lebensweise:
- Wenn manche oberdevonische Tetrapoden sekundär wasserlebend waren, müßte es viele landangepaßte Tetrapoden im Devon gegeben haben; doch davon fehlt jede Spur (Clack 2002a, 135).
- Das Rückkehr-Szenario ist unplausibel, da die zunächst ans Landleben angepaßten Formen bei ihrer Rückkehr auf harte Konkurrenz stoßen müßten (s. u.: „Selektionsdrücke“). Es wird oft argumentiert, daß Konkurrenz die Fische aufs Land getrieben hätte, wo sie einen konkurrenzfreien Raum hatten. Vor diesem Hintergrund erscheint eine Rückkehr ins Wasser nicht glaubhaft.
Dilemma.
Evolutionstheoretisch tut sich hier ein Dilemma auf: Sowohl die Entstehung der Tetrapoden-Merkmale im Wasser als auch die Rückkehr ins Wasser sind unplausibel. Insgesamt heißt das: es gibt kein plausibles Szenario für die evolutive Entstehung der oberdevonischen Tetrapoden. Man kann daraus schließen, daß die oberdevonischen Formen, die möglicherweise in einem vegetationsreichen Uferbereich als lauernde Räuber gelebt haben, für diesen speziellen (oder einen ähnlichen) Lebensraum konstruiert waren. Dies läßt sich im Rahmen eines Schöpfungsmodells zwanglos verstehen. In Uferbereichen dürfte es eine Vielfalt an ökologischen Nischen gegeben haben, darunter möglicherweise auch (damalige) Lebensräume, die es in dieser Form heute nicht mehr gibt. Dieser Vielfalt könnte eine anatomische Formenfülle mit unterschiedlichen Konstruktions-Mosaiken des Kiemenapparats, der Extremitäten, der Wirbelsäule, des Schwanzes usw. entsprechen.
Selektionsdrücke – oder: Warum entstanden die Tetrapoden?
|
Die Abstammung der heute lebenden Amphibien (Froschlurche, Schwanzlurche und Blindwühlen, als Lissamphibia zusammengefaßt) ist ungeklärt. Die fossilen paläozoischen Formen sind nämlich in vieler Hinsicht so „hochentwickelt“, daß man die Frösche und Schwanzlurche davon nicht ableiten kann. Carroll (1993, S. 196) weist darauf hin, daß es eine „überraschende Lücke“ in der Fossilüberlieferung zwischen frühen Amphibien einerseits und Frosch- und Schwanzlurchen sowie Blindwühlen andererseits gibt. „Wenn sie im Jura zuerst im Fossilbeleg erscheinen, sehen sowohl die Frösche als auch die Salamander in der Skelettanatomie im Prinzip modern aus.“ Auch während der gesamten Fossilüberlieferung unterscheiden sich diese Gruppen stark voneinander.
|
|
Clack (2002a, 99) bezeichnet viele Szenarien zur Entstehung der Tetrapoden als „Mythologie“. Die Vorstellung, daß solche Fische, die auch Lungen besaßen, prädestiniert fürs Überleben waren, wird aus zwei Gründen in Frage gestellt: 1. Viele Fleischflosser-Fische überlebten die weltweiten Aussterbeereignisse im Oberdevon. So „nötig“ war der Schritt aufs Land daher gar nicht; ein entsprechender Selektionsdruck ist also wenig plausibel. 2. Wahrscheinlich hatten auch die frühen Strahlflosser-Fische Lungen, so daß der Besitz von Lungen wohl keine entscheidende Prädisposition für das Landleben darstellt (Clack 2002a, 100).
Auf Alfred Romer geht eine sehr populär gewordene Vorstellung zurück, die man mit etwas Humor als „Von Tümpel zu Tümpel sich schleppen müssen-Hypothese“ bezeichnen könnte. Demnach soll die Tetrapodenextremität dadurch entstanden sein, daß devonische Quastenflosser in austrocknenden Tümpeln nur dadurch dem Tod entrinnen konnten, daß sie sich zu einem nahegelegenen Tümpel schleppen konnten. In der Old Red-Ausbildung (Red Beds) des Devons in Nordeuropa und Nordamerika sind z. B. durch Trockenrisse in den Oberflächen der roten Sandsteine zumindest kurzzeitige Austrocknungsepisoden belegt. Das wird als Hinweis auf ein trockeneres Klima gewertet. Für das Erreichen nahegelegener Tümpel waren kräftige, fleischige Flossen sehr nützlich, und entsprechend soll sich ein Selektionsdruck zur Entwicklung von Beinen aufgebaut haben. Das Romersche Szenario ist ausgesprochen paradox: Die Beine sollen sich entwickelt haben, damit die zu Verbeinern werdenden Fische im Wasser bleiben konnten – ein solches Konzept ist also schon in sich nicht schlüssig (Coates & Clack 1995, 375).
Dennoch hat es große Popularität gewonnen und findet sich in fast jedem Lehrbuch zur Evolution der Lebewesen. Die Romersche Hypothese wird heute jedoch aus weiteren Gründen als sehr fragwürdig angesehen, z. B. werden heute die Red Beds des Devons nicht mehr unbedingt mit trockenem Klima in Verbindung gebracht. Zudem ist nicht gesichert, daß in den Gebieten, in welchen die Tetrapoden entstanden, ein trockenes Klima herrschte. Viele devonische Sedimente sind jedenfalls Fluß- oder Küstensedimente (Clack 2002a, 101). Die ökologischen Bedingungen, unter denen die frühen Tetrapoden entstanden, sind nicht bekannt; es könnten statt Süßwasser- auch Brackwasserbedingungen gewesen sein. Im Oberdevon gab es offenbar eine große Vielfalt an Biotopen. Auch von daher spielen Trockengebiete im Devon für die Entstehung der Tetrapoden möglicherweise keine Rolle (Clack 2002a, 102). Zudem entspricht keine der frühen Tetrapoden-Umwelten, wie sie aus der Sedimentausbildung (Fazies) erschlossen wird, in denen die Tetrapoden gefunden werden, dem Romerschen Szenario der saisonalen Trockenzeiten (Coates & Clack 1995, 381).
Dazu kommt noch folgende Beobachtung: Moderne Amphibien suchen bei Austrocknung keine neuen Wasserlöcher, sondern sammeln sich in den austrocknenden Pools und sterben oft. Dieses Verhalten wird auch bei bestimmten Massenfunden fossiler Reptilien angenommen (z. B. Müller 1992, 350f.). Ein Selektionsdruck würde Schutz gegen Austrocknung fördern, nicht den Erwerb von (längeren) Beinen. Im Devon wäre das wohl kaum anders gewesen (Clack 2002a, 100). Die heutigen Lungenfische und einige primitive Strahlflosser-Fische besiedeln Biotope, die zeitweise austrocknen. Lungenfische versuchen jedoch nicht, das Land zu erobern, sondern machen einen Sommerschlaf in einem selbstgegrabenen Schacht, in dem sie sich mit einer gegen Austrocknung schützenden Kapsel aus Schleim und Erdreich umgeben. Dabei wird der Stoffwechsel drastisch heruntergefahren (Bray 1985, 310; Clack 2002a, 101).
Dazu kommen die neueren Hinweise, daß die Tetrapodenextremität im Wasser entstanden sein dürfte (s. o.). Damit ist das Romersche Konzept wohl endgültig aus dem Rennen.
Doch auch das Szenario der Entstehung von Beinen im Wasser erscheint aus Selektionsgründen nicht plausibel. Denn weshalb sollten im Wasser aus Flossen Beine mit Fingern entstehen? Müßte Selektion das nicht verhindern, da es für die Fähigkeit zum Schwimmen eine Einschränkung bedeutet? Extremitäten mit Fingern ermöglichen zwar ein besseres Kraulen in dichter Vegetation, gleichzeitig wird aber ein bewährtes Organ (Flossen) aufgegeben, obwohl sich der Lebensraum nicht wesentlich gewandelt hat. Immerhin erscheint ein solches Szenario plausibler als die Romersche Hypothese. Sie macht jedoch allenfalls den Erwerb der Finger durch die Selektionsbedingungen plausibel, nicht den Schritt ans Land. (Und selbst wenn selektiv nichts gegen eine evolutive Entstehung der Tetrapodenextremität im Wasser sprechen würde, wäre nur eine notwendige und keine hinreichende Bedingung für die evolutive Entstehung erfüllt.)
Die Rückkehr ins Wasser ist von den Selektionsbedingungen her noch weniger schlüssig. Gerade erst der Konkurrenz im Wasser entflohen, mühsam an ein teilweises Landleben angepaßt, dabei Einrichtungen fürs Leben im Wasser verloren: in dieser Situation sollen frühe Tetrapoden „fit“ gewesen sein für die erneute Konkurrenz im Wasser?
Als weitere Idee zur Entstehung von Beinen wurde die Vorstellung geäußert, sie wären entwickelt worden, um dem Tier die Möglichkeit zum Eingraben im Schlamm zu geben. Diesem Szenario steht aber entgegen, daß heutige Formen zum Eingraben nicht die Beine benutzen (Clack 2002a, 100).
Obwohl der Lungenfisch Neoceratodus am ehesten taugliche Beine für eine Fortbewegung an Land besitzt, lebt er – verglichen mit anderen heutigen Lungenfischen – ausgerechnet am meisten im Wasser (Edwards 1989, 246). Protopterus benutzt seine Flossen, um sich auf dem Boden unter Wasser fortzubewegen. So war es nach der Vermutung von Edwards (1989, 246) wohl auch bei den Osteolepiformes (den mutmaßlichen Tetrapoden-Vorläufern). Außerdem: Sollten die Beine zu Grabwerkzeugen evolviert sein, wären sie nicht zum Laufen geeignet gewesen (Clack 2002a, 101; unter Bezugnahme auf eine Arbeit von Ewer 1955).
 |
| Keine der vorgetragenen Hypothesen zum Ursprung der Tetrapoden-extremität ist derzeit testbar. |
 |
|
Die Fortpflanzungsweise der frühen Tetrapoden ist unbekannt. Selektionsdrücke in diesem Gebiet sind somit spekulativ (Clack 2002a, 101). Ein anderer möglicher Selektionsfaktor könnte Populationsdruck bzw. Feinddruck sein (Clack 2002a, 101, 104), doch auch dies ist spekulativ. Keine der vorgetragenen Hypothesen zum Ursprung der Tetrapodenextremität ist derzeit testbar (Clack 2002a, 136). Zu Selektionsdrücken und Lebensbedingungen der oberdevonischen Tetrapoden gibt es zwar zahlreiche Ideen, die Evidenzen sind jedoch unklar. Gegen die meisten dieser Ideen können plausible Gegenargumente formuliert werden. Ohne zusätzliche Fossilfunde sind derzeit nur Vermutungen möglich (Clack 2002a, 104).
Weitere Probleme.
Zu den genannten Punkten kommen noch folgende Beobachtungen erschwerend hinzu:
- Heutige Fische zeigen, daß es keine logische Verknüpfung zwischen dem Verlust von Flossenstrahlen und Flossennetzen, Bildung von Fingern und Exkursionen aufs Land gibt. So sind Aale diejenigen Fische, die heute die längsten Wanderungen auf dem Land unternehmen; deren paarige Flossen sind aber stark reduziert (Clack 2002a, 102).
- Es gibt moderne Strahlflosser mit Anpassungen ans Landleben und solche, die fingerartige Strukturen von Flossenstrahlen ausgebildet haben. Diese zwei Gruppen überlappen sich nahezu nicht. Fische mit Fingern gehen nicht an Land, und solche, die aufs Trockene gehen, haben keine fingerartigen Strukturen an ihren Extremitäten (Clack 2002a, 103). Der in der Tiefsee lebende Sargassum-Anglerfisch (Histrio histrio) hat z.B. acht Finger. Er benutzt seine beinartigen Flossen wie Tetrapoden, um auf dem Meeresboden zu laufen und sich so an Beute heranzuschleichen (Edwards 1989, 249ff.). Die große Zahl analoger Verhältnisse beim Unterwassergehen von Anglerfischen stützt die These, daß die Tetrapoden-Extremität unter Wasser entstanden ist. Ein Zusammenhang zwischen der Ausbildung beinartiger Extremitäten und Landleben ist unter heutigen Formen jedoch nicht erkennbar (Clack 2002a, 103).
 |
| Abb. 23: Der in der Tiefsee lebende Anglerfisch Histrio mit Fingern an den Flossen. (Nach Clack 2002a) |
|
Damit stellt sich aber die Frage, ob mit dem Erwerb von Fingern unter Wasser überhaupt Argumente für den Schritt an Land gewonnen sind. Offenbar haben die meisten gefingerten Formen diesen Schritt nicht getan.
- Die Strahlflosser-Fische werden bekanntlich in keine nähere Verbindung mit dem Ursprung der Tetrapoden gebracht. Dennoch sind unter ihnen 59 amphibisch lebende Arten aus 16 marinen und limnischen Familien bekannt. Keine davon hat Analoga zu den Tetrapoden-Beinen entwickelt (Coates & Clack 1995, 301). Bei diesen Formen wäre ein Selektionsdruck in diese Richtung am ehesten nachvollziehbar.
Aus diesen Befunden ergibt sich als Gesamtergebnis: Ein Zusammenhang zwischen der Eroberung des Landes (bzw. einer amphibischen Lebensweise) und dem Erwerb der Vierbeinigkeit mit Fingern ist durch Daten wenig begründet.
Vielfingrigkeit – ein Primitivmerkmal?
Unter den ältesten Tetrapoden besitzen einige Gattungen mehr als fünf Finger: Acanthostega hat acht Finger, Tulerpeton vorne sechs und hinten sieben, Ichthyostega hinten sieben (was allerdings von Jarvik [1996] bestritten wird; die Verhältnisse bei der Vorderextremität sind unbekannt). Alle karbonischen Tetrapoden haben fünf Finger (oder weniger). Der Besitz von mehr als fünf Fingern (Polydaktylie) erscheint somit als Indiz dafür, daß noch keine „festen evolutionären Bahnen“ erreicht seien. Das sei typisch für frühe Phasen der Evolution neuer Baupläne. Erst mit der Fünffingrigkeit seien dagegen stabile Verhältnisse erreicht. Diese Deutung einer instabilen Frühphase ist jedoch kaum prüfbar; dazu müßten die genetischen Verhältnisse bekannt sein, was bei ausschließlich fossil erhaltenen Organismen nicht möglich ist.
Ein im Jahr 2003 beschriebenes Tetrapoden-Fossil aus der Trias läßt zudem Zweifel an dieser Deutung aufkommen. Dieses bislang noch unbenannte marine Reptil hat an der Vorderextremität vor dem ersten Finger einen bzw. zwei zusätzliche gut entwickelte Finger (Wu et al. 2003). Die Morphologie der Hand und des Fußes besitzt eine starke Ähnlichkeit mit den Gliedmaßen der frühesten Tetrapoden, stellen Wu et al. (2003) fest. Es handelt sich um den ersten Fall einer Polydaktylie bei einem jüngeren Fossil, die den Verhältnissen der oberdevonischen Tetrapoden gleicht. Die Autoren sprechen von einem schlagenden Beispiel für Konvergenz.
 |
| Abb. 24: Vielfingrigkeit bei einem (noch unbenannten) Tetrapoden aus der Trias (SSTM 5025), Jialingjiang-Formation, Hubei-Provinz, China. Dieses Fossil stellt in Frage, daß Vielfingrigkeit ein Primitivmerkmal ist. (© X.C. Wu; Abdruck mit freundlicher Genehmigung) |
|
Polydaktylie kommt auch bei den meisten Fischsauriern vor. Sie ist allerdings von deutlich anderer Art als bei den devonischen Tetrapoden. Die Fischsaurier zeigen jedoch ebenso wie das neubeschriebene vielfingrige Trias-Fossil: Eine strenge Fixierung auf maximal fünf Finger ist unter evolutionstheoretischer Perspektive eben nicht gegeben. Daß nach einer instabilen evolutionären Frühphase eine Fixierung erfolgt sein soll, ist daher eine fragwürdige Deutung. Es dürfte ein fruchtbarer heuristischer Ansatz sein, auch bei den Oberdevon-Formen nach der Funktionalität der Polydaktylie zu fragen, anstatt diese als zufälliges evolutionäres Durchgangsstadium zu interpretieren.
Evolutions- oder Besiedlungsabfolgen?
Wie gezeigt wurde, ist es kaum möglich, aus der Vielfalt devonischer Formen evolutionäre Abfolgen herauszudestillieren. Das gilt in noch viel größerem Maße für die Tetrapoden des Karbons. Zu den genannten Schwierigkeiten gesellen sich noch zwei weitere Problempunkte evolutionärer Rekonstruktionen.
- Es ist nicht sicher, vielleicht nicht einmal wahrscheinlich, daß die fossil erhaltenen Tetrapoden repräsentativ sind für die jeweiligen Zeiten, denen die Fundschichten zugeordnet werden. Denn wasserlebende Formen werden gegenüber den landlebenden bevorzugt fossil überliefert (Smithson et al. 1994, 383; Milner et al. 1986). Die bekannten Fossilien belegen nur eine sehr eingeschränkte Anzahl an Lebensräumen, und zwar bevorzugt wassernahe. Bolt & Lombard (1992, 380) vermerken daher, daß manche Organismen immer verborgen bleiben werden. In bergigen Gebieten entstehen fast keine dauerhaften Sedimente, in denen Fossilien überliefert werden könnten. Deshalb weiß man nur wenig über die mehr ans Landleben angepaßten Tetrapoden jener Zeit (Clack 2002a, 195f.). Vielleicht spiegeln die Fossilien teilweise ökologische Zonierungen anstelle evolutionärer Abfolgen wider.
- Die Fossilfunde stammen insgesamt großenteils aus einer relativ begrenzten geographischen Region. Lange Zeit waren karbonische Tetrapoden – abgesehen von einer Handvoll weniger interessanter Formen – nur von der Südküste Euroamerikas bekannt (Milner et al. 1986, 12). Diese Autoren werfen die Frage auf: Repräsentieren die Abfolgen dieser fossil überlieferten Tiere eine evolutionäre Sequenz oder nur eine lokale Sukzession von Lebensgemeinschaften, die jeweils durch Einwanderer aus anderen Gegenden abgelöst wurden, oder handelt es sich vielleicht um eine komplexe Mischung aus beidem?
Mittlerweile wurde eine diverse Tetrapodenfauna in der Südhemisphäre (Australien) aus dem höheren Unterkarbon (mittleres Viséum) entdeckt, was eine neue Situation darstellt (Thulborn et al. 1996). Zusammen mit vielen Süßwasserfischen sind mindestens drei Tetrapoden-Typen gefunden worden. Die Verbreitung ist geographisch so ausgedehnt, daß an eine mehrfach unabhängige Entwicklung von Tetrapodengruppen gedacht wird (Milner 1996, 742). Die Möglichkeit, daß fossile Abfolgen auf Einwanderungswellen in passende neue Lebensräume anstatt auf evolutionäre Tendenzen zurückgehen, bleibt aber weiter offen.
|
|
Zusammenfassung
- Die evolutionstheoretischen Vorstellungen über die oberdevonischen Tetrapoden und mögliche Fisch-Vorläufer haben sich in den letzten Jahren stark gewandelt. Die Vierbeinigkeit wurde bis vor kurzem als gleichbedeutend mit einer (teilweise) landlebenden Lebensweise gleichgesetzt. Dies trifft nach heutigen Erkenntnissen nicht mehr zu. Die Vierbeiner des Oberdevons lebten hauptsächlich oder sogar ausschließlich im Wasser. Diese Einschätzung trifft neuerdings auch auf das berühmte „erste Amphib“ Ichthyostega zu, das diesen Status damit verloren haben dürfte.
- Möglicherweise haben die oberdevonischen Tetrapoden mit der Eroberung des Landes nichts zu tun (auch nicht in evolutionstheoretischer Perspektive). Sie sind alle mehr oder weniger deutlich spezialisiert (vermutlich auf eine Lebensweise in vegetationsreichen Uferzonen) und eignen sich nicht als direkte Vorläufer karbonischer Formen. Es ist durchaus denkbar, daß es sich bei den oberdevonischen Tetrapoden – gemäß evolutionärer Interpretation – um eine Sackgasse handelt, die nicht aufs Land führte, sondern der Weg an Land über andere, unbekannte Formen ging.
- Die Ausbildung von Fingern muß nicht in einem Zusammenhang mit der Eroberung des Landes stehen, wie auch das Vorkommen von Fingern bei Sauripterus beweist, der nicht näher mit Tetrapoden verwandt ist. Aber die Vierbeinigkeit stellt für die Eroberung des Landes eine bessere Präadaptation dar als der Besitz von Flossen.
Erst recht müssen viele andere Tetrapodenmerkmale bei Fischen nichts mit einem Übergang zum Landleben zu tun haben. Dies wird besonders bei Acanthostega deutlich. Diese Gattung weist zu 2/3 Tetrapodenmerkmale auf (Clack & Coates 1995, 359), ist aber nach Einschätzung der Paläontologen vollständig wasserlebend.
- Der Übergang einzelner Schlüsselmerkmale ist schlecht oder gar nicht fossil belegt. Der Erwerb von Fingern ist nicht durch Fossilfunde dokumentiert, ebensowenig wie der Umbau der Ohrregion zum Hören außerhalb des Wassers. Der Übergang von paddelförmigen Schwimmbeinen zu solchen, die Körpergewicht an Land tragen können und für ein Gehen an Land geeignet sind, ist ebenfalls nicht überzeugend fossil belegt. Erwähnung kann hier auch der schnelle (d. h. fossil nicht belegte) Umbau des Hirnschädels finden, ebenso der Bau des Beckengürtels, der auch bei den ältesten bekannten Tetrapoden sehr verschieden ist vom Bau der betreffenden Strukturen bei Fischen, wie Clack (2002a, 42) feststellt.
Andererseits zeigen viele Merkmale durchaus graduelle Abstufungen bei einzelnen Gattungen, etwa in der Anzahl oder Größe von Knochen im Kopf-/Hals-/Schultergürtel-Bereich. Man muß aber nicht davon ausgehen, daß diese unterschiedlichen Ausprägungen funktionsloser Ausdruck von evolutionären Umbauten darstellen; vielmehr waren sie für die jeweilige Lebensweise passend. (Dies führt in das Thema der Rudimentation, das hier nicht erörtert werden kann; siehe dazu Junker 2002a.) Der Vielseitigkeit der Lebensräume in Uferbereichen könnte eine graduelle Vielfalt von Merkmalsausprägungen entsprechen. Dieser Befund der graduellen Unterschiede gibt evolutionären Deutungen eine gewisse Plausibilität, allerdings müssen vielfach Konvergenzen angenommen werden (s.u.).3
- Aufsteigend in der Schichtenfolge kann eine vergröberte Betrachtungsweise eine Zunahme an tetrapodenartigen Merkmalen feststellen. Im oberen Oberdevon (Famennium) gibt es erste Tetrapoden, die im unteren Oberdevon (Frasnium) fehlen (oder als solche nicht eindeutig fossil dokumentiert sind). Im Unterkarbon treten unzweifelhaft landlebende Formen auf, die im Oberdevon nicht überliefert sind. Im Detail betrachtet passen mögliche evolutionäre Abfolgen jedoch oft nicht zum Fundniveau in der Schichtenfolge. So ist der „tetrapodenähnlichste Fisch“ Panderichthys älter als Eusthenopteron. Ventastega gilt als einer der primitivsten oberdevonischen Tetrapoden, obgleich er im Oberdevon zu den jüngsten Vierbeinern gehört. Im Unterkarbon treten ausgerechnet die höchstspezialisierten Formen, die beinlosen Aïstopoda, zuerst in der Fossilüberlieferung auf.
- Nach der „Romer-Lücke“ tauchen die Vierbeiner des höheren Unterkarbons plötzlich in großer Vielfalt auf. Die Formenvielfalt ist so groß, daß es kaum möglich ist, klare Verwandtschaftsbeziehungen zwischen den einzelnen Gattungen herzustellen. Dendrogramme offenbaren zahlreiche Konvergenzen und Rückentwicklungen.
- Die Selektionsdrücke, die zur Entstehung der Vierbeinigkeit bei den devonischen Formen führten, sind unklar. Es gibt starke Argumente für eine Entstehung von Fingern und Tetrapodenextremität im Wasser, aber ähnlich viel spricht dagegen bzw. man kann Argumente dafür nennen, daß die oberdevonischen Tetrapoden sekundär wasserlebend sind. Die jeweiligen Argumentationsstrategien schließen die jeweils andere Position aus. Dieses Dilemma ist evolutionstheoretisch ungelöst.
- Wie alternativ skizziert wurde, könnten die oberdevonischen Formenabfolgen ökologische Zonierungen statt eines evolutionstheoretischen Szenarios wiederspiegeln.
|
Anmerkungen
1 Als Beleg seien hier einige Zitate von Paläontologen angeführt: „... many characters show conflicts (incongruence) in their distribution. In other words, the distribution of primitive and derived characters do not always overlap cleanly and simply in the way one might expect“ (Clack 2002a, 14). „It has long been clear that none of the early tetrapod groups are ideal ancestors for any of the rest. All exhibit a mosaic of primitive and derived characters that indicate significant periods of independent evolution since the initial divergence of each group“ (Carroll 1992, 49). Carroll (1998, 144) stellt daher allgemein fest: „There is currently no consensus as to the specific interrelationships of any of the major groups of Palaeozoic tetrapods“ (Carroll 1998, 144; Hervorhebung nicht im Original). Weitere ähnliche Zitate könnten angeführt werden.
2 In jüngster Zeit fassen Schoch & Carroll (2003, 315) die Situation wie folgt zusammen: „Recent phylogenetic analyses by Clack (2002a) and Clack & Ahlberg (in press), based on newly recognized species from the Lower Carboniferous, show very poor resolution of sister-group relationships among Paleozoic tetrapods. Tetrapods from the Upper Devonian, Lower Carboniferous, and Upper Carboniferous appear as a succession of radiations, each of which are characterized by unresolved polytomies“ (Hervorhebung nicht im Original).
3 Ein Beispiel dazu: Ahlberg & Clack (1998) präsentieren eine cladistische Analyse zu den Kiefern früher Tetrapoden. Sie können zwar eine Abfolge von Gattungen beschreiben, bemerken aber: „What we cannot always say is which of these events happened several times in parallel, or whether there were many instances of character reversal“ (Ahlberg & Clack 1998, 38). The distribution of lateral line characters [am Kiefer] is not wholly congruent, and there may have been a considerable amount of parallelism and reversal in this process. The earliest known example of the fully derived condition is ‘Tulerpeton’“ (Ahlberg & Clack 1998, 38). „There is evidently a fair amount of incongruence in the details of the dentition, but the outlines of the general trends are clear“ (Ahlberg & Clack 1998, 40).
|
Literatur
- Ahlberg PE & Clack JA (1998)
- Lower jaws, lower-tetrapods – a review based on the Devonian genus Acanthostega. Trans. R. Soc. Edinb. 89, 11-46.
- Bolt JR & Lombard RE (1992)
- The nature and quality of the fossil evidence of otic evolution in early tetrapods. In: Webster D, Fay RR & Popper AN (eds) The Evolutionary Biology of Hearing. New York: Springer Verlag, pp 377-407.
- Bray AA (1985)
- The evolution of the terrestrial vertebrates: environmental and physiological considerations. Phil. Trans. R. Soc. Lond. 309, 289-322.
- Carroll RL (1982)
- Early evolution of reptiles. Annu. Rev. Ecol. Syst. 13, 87-109.
- Carroll RL (1992)
- The primary radiation of terrestrial vertebrates. Annu. Rev. Earth Planet. Sci. 20, 45-84.
- Carroll RL (1998)
- Cranial anatomy of ophiderpetontid aistopods: Palaeozoic limbless amphibians. Zool. J. Linn. Soc. 122, 143-166.
- Clack JA (1994)
- Silvanerpeton miripedes, a new anthracosauroid from the Viséan of East Kirkton, West Lothian, Scotland. Trans. R. Soc. Edinb. Earth Sci. 84, 369-376.
- Clack JA (2002a)
- Gaining Ground. The origin and evolution of Tetrapods. Bloomington and Indianapolis.
- Clack JA (2002b)
- An early tetrapod from ‘Romer’s Gap’. Nature 418, 72-76.
- Coates MI & Clack JA (1995)
- Romer’s gap: tetrapod origins and terrestriality. In: Arsenault M, Lelièvre H & Janvier P (eds) Studies on early vertebrates (VIIth International Symposium, Miguasha Parc, Quebec). Paris: Bull. Mus. Natl. Hist. Nat. 17, 373-388.
- Daeschler EB (2000)
- Early tetrapod jaws from the Late Devonian of Pennsylvania, USA. J. Paleont. 74, 301-308.
- Edwards JL (1989)
- Two perspectives on the evolution of the Tetrapod limb. Am. Zool. 29, 235-254.
- Janis CM & Farmer C (1999)
- Proposed habitats of early tetrapods: gills, kidneys, and the water-land transition. Zool. J. Linn. Soc. 126, 117-126.
- Jarvik E (1996)
- The Devonian tetrapod Ichthyostega. Fossils Strata 40, 1-206.
- Junker R (2002)
- Ähnlichkeiten, Rudimente, Atavismen. Studium Integrale. Holzgerlingen.
- Milner A (1996)
- Early amphibian globetrotters? Nature 381, 741-742.
- Milner AR, Smithson TR, Milner AC, Coates MI & Rolfe WDI (1986)
- The search for early tetrapods. Modern Geology 10, 1-28.
- Müller AH (1992)
- Lehrbuch der Paläozoologie, Bd. I. Allgemeine Grundlagen. Jena; Stuttgart.
- Schoch RR & Carroll RL (2003)
- Ontogenetic evidence for the Paleozoic ancestry of salamanders. Evol. Dev. 5, 314-324.
- Smithson TR, Carroll RL et al. (1994)
- Westlothiana lizziae from the Viséan of East Kirkton, West Lothian, Scotland. Trans. R. Soc. Edinb. Earth Sci. 84, 383-412.
- Thulborn T, Warren A, Turner S & Hamley T (1996)
- Early Carboniferous tetrapods in Australia. Nature 381, 777-780.
- Westenberg K (1999)
- From fins to feet. Nat. Geogr. 195, 114-126.
- Wu X-C, Li Z, Zhou B-C & Dong Z-M (2003)
- A polydactylous amniote from the Triassic period. Nature 426, 516.
|
|
|  |
