|
Komplexe Pflanzen überraschend früh
von Reinhard Junker
Studium Integrale Journal
10. Jahrgang / Heft 2 - Oktober 2003
Seite 74 - 76
|
 |
Frühe Landpflanzen mit komplexen Merkmalen sind in deutlich älteren Schichten gefunden worden als vergleichbare bisher bekannte Pflanzengattungen. Sie sind komplexer als die primitivsten bekannten Land-Gefäßpflanzen.
| |
|
|
Abb. 1: Macivera gracilis, von Kotyk et al. zu den Zosterophyllophyten gestellt, ist aber als Mosaikform zu betrachten. Die Pflanze besaß nackte, dünne, gabelig verzweigte Stengel. Da die Sporangien in kleinen Gruppen am Ende der Achse (wenn auch nicht in Reihen) angeordnet und direkt am Stengel sitzend waren, stellen Kotyk et al. (2002) sie zu den Zosterophyllophyten (s. Kasten). Dazu paßt auch der für diese Gruppe typische spezialisierte Querspalt der Sporangienöffnung. Allerdings waren die Sporangien länglich, waren wahrscheinlich noch unreif (denn die darin enthaltenen Sporen waren noch nicht ausgereift), und der Sporangienstand daher im ausgereiften Zustand vermutlich lockerer. Diese Merkmale sind typisch für Rhyniophyten. Es handelte sich also um eine Mosaikform. (Nach Kotyk et al. 2002) |
Über „morphologisch komplexe“ Fossilien früher Landpflanzen aus dem Obersilur der Insel Bathurst in Kanada berichten KOTYK et al. (2002). Die Funde sind bedeutsam, weil es sich hauptsächlich um Arten handelt, die bisher erst aus dem Unterdevon bekannt waren und evolutionär als deutlich „fortschrittlicher“ eingeschätzt werden als die einfachsten bekannten fossilen Landpflanzen. Das heißt: Sehr früh in der Fossildokumentation eindeutiger Landpflanzen treten bereits verschiedene abgeleitete („höherentwickelte“) Formen auf. (Zur Stratigraphie vgl. Abb. 1.)
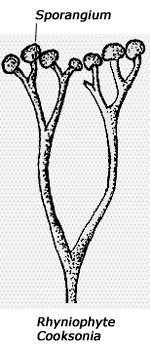
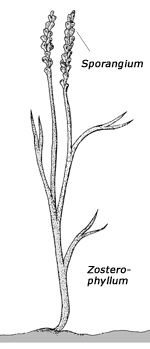
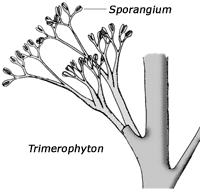
Die ersten Land-Gefäßpflanzen – früher als „Psilophyten“ (= „kahle Pflanzen“) bezeichnet – wurden seit der Revision der Klassifikation durch BANKS (1975) in drei Gruppen unterteilt, die alle blattlos sind: Rhyniophyten, Zosterophyllophyten und Trimerophyten (siehe Kasten). (Die Grenzen zwischen diesen Gruppen sind durch zahlreiche mittlerweile entdeckte Mosaikformen inzwischen teilweise etwas verwischt; vgl. BANKS 1992; KENRICK & CRANE 1997.) Dabei gelten die am einfachsten gebauten Rhyniophyten als die ursprünglichsten Formen, während die anderen beiden Gruppen als höherentwickelt eingestuft werden. Bislang waren diese Pflanzen erst ab dem Unterdevon fossil bekannt – mit Ausnahme von meist sehr bruchstückhaft erhaltenen Fossilien, die in die Nähe der Rhyniophyten gestellt werden und wegen unsicherer systematischer Stellung und unbekannter anatomischer Verhältnisse von manchen Autoren als Rhyniophytoiden zusammengefaßt werden. Rhyniophytoiden sind bereits ab dem mittleren Silur fossil dokumentiert. Darüber hinaus gehören zu den frühesten Landpflanzen aber auch mit Mikrophyllen (schmalen Blättchen) ausgestattete Formen, die zu den Bärlapppflanzen gerechnet werden (vgl. Abb. 3).
|
Rhyniophyten: Blattlos und ohne Auswüchse; Luftsprosse meist gabelig verzweigt; Sporangien meist endständig und einzeln, rundlich bis länglich; Sproß mit dünnem zentralem Xylem (Protostele), wobei das Protoxylem (die ersten sich bildenden Leitgefäße) sich in der Mitte befindet.
Zosterophyllophyten: Blattlos; gabelig oder pseudomonopodial verzweigt (d. h. mit scheinbarer Hauptachse); Sporangien nierenförmig bis rundlich, seitenständig, oft in Ähren, Öffnungsmechanismus (zum Entlassen der Sporen) mit Querspalt; Xylem groß, elliptisch oder rund, Protoxylem am Rand.
Trimerophyten: Blattlos; Stengel meist mit Hauptachse (pseudomonopodial), mit spiralig angeordneten und gabelig abgehenden Zweigen; Seitenzweige gabelig oder dreiteilig verzweigt; fertile Zweige (vielfach verzweigt) enden in zahleichen paarigen, spindelförmig-elliptischen Sporangien; fertile und sterile Regionen sind deutlich unterschieden; Sporangien öffnen sich der Länge nach; die Stengel sind glatt oder stachelig; das Xylem ist groß und rund, elliptisch oder fast viereckig, das Protoxylem befindet sich in der Mitte.
|
Die Funde, über die nun KOTYK et al. (2002) berichten, gehören hauptsächlich den Zosterophyllophyten (vgl. Kasten) an, und unter diesen sind auch komplexere Formen vertreten. Die Autoren beschreiben Arten aus den Gattungen Zosterophyllum, Bathurstia und Distichophyton mit ausgeprägt ährenförmigem Sporangienstand – ein Merkmal, das als evolutionär abgeleitet eingestuft wird (Sporangien = Sporenbehälter; vgl. Abbildungen im Kasten). Neu beschrieben wird die Gattung Macivera, welche die Autoren ebenfalls zu den Zosterophyllophyten stellen – eigentlich handelt es sich um eine Mosaikform aus rhyniophytischen und zosterophyllophytischen Merkmalen (siehe Abb. 1 und dortigen Bildtext). Nur eine fragmentarisch erhaltene Art kann den Rhyniophytoiden zugeordnet werden.
 |
| Abb. 2: Stratigraphische Unterteilung des Silurs und des Unterdevons. Die hier beschriebenen neuen Funde stammen aus dem oberen Ludlow (Obersilur). |
|
KOTYK et al. (2002, 1004) halten die Flora der Bathurst-Insel für die derzeit bestdokumentierte Flora von silurischen Landpflanzen mit erheblicher Vielfalt und Komplexität im Bau der Pflanzen. Das frühe Auftreten läßt für die Autoren folgende Fragen aufkommen: Ist die gruppenweise oder sitzende Anordnung der Sporangien primitiv bei den Zosterophyllophyten? (Bislang wurde dies verneint.) Bieten diese Merkmale einen Selektionsvorteil? Können diese Merkmale in einem Zusammenhang mit biotischen oder abiotischen Faktoren gesehen werden? Das Gruppieren von Sporangien könnte einen gewissen Schutz vor Austrocknung bieten, doch stehen funktionelle Betrachtungen auf unsicherem Boden.
Nur zwei andere silurische Fundorte mit Pflanzen vergleichbarer Komplexität waren bisher beschrieben: Die Baragwanathia-Flora Australiens (Obersilur) und die rätselhafte Gattung Pinnatiramosus aus dem Untersilur Chinas. Baragwanathia (Abb. 3) wird zu den Bärlappgewächsen gestellt, die unter den frühesten Landpflanzen wegen ihrer Mikrophylle und stengelständigen Sporangien als relativ hochentwickelt gelten. Die Datierung der Baragwanathia-Flora ist allerdings seit langem umstritten, ohne daß eine endgültige Lösung gefunden wurde. Manche Autoren datieren sie ins Unterdevon. Die silurische Datierung erfolgt aufgrund von Leitfossilien. (Einstufungen mit Leitfossilien gelten aber sonst allgemein als sehr zuverlässig, und man fragt sich, ob das hier nicht deshalb zum Teil angezweifelt wird, weil die „Entwicklungshöhe“ dieser Pflanzen evolutionär „unpassend“ ist.)
fertil: fruchtbar.
Mikrophylle: kleine, meist ungegliederte, schmale Blätter.
Protoxylem: die ersten, noch dünnen wasserleitenden Gefäße, die im Stengel gebildet werden.
Sporangien: Sporenbehälter, im Umriß meist rundlich, elliptisch oder nierenförmig. Sporen sind einzellige, ungeschlechtliche Fortpflanzungskörper.
Xylem: Gefäß- oder Holzteil, wasserleitender Teil des Leitbündels.
|
Die Natur der untersilurischen (!) Gattung Pinnatiramosus ist umstritten, weil nicht ganz klar ist, ob es sich überhaupt um eine Landpflanze handelt. CAI et al. (1996) haben allerdings starke Indizien für diese Interpretation vorgelegt.
|
|
|
Schlußfolgerungen
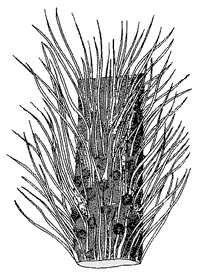 |
| Abb. 3: Das älteste fossil dokumentierte Bärlappgewächs mit dem wohlklingenden Namen Baragwanathia. (Nach Stewart & Rothwell 1993). |
|
Mit den neuen Funden ist in mehrerer Hinsicht eine neue Lage in der Beurteilung evolutionärer Vorstellungen über das Auftreten der ersten Landpflanzen eingetreten:
Das frühe fossile Auftreten der Baragwanathia-Flora erscheint nicht mehr so ungewöhnlich, wie KOTYK et al. (2002, 1012) bemerken. Vielleicht wird das den Streit um die Datierung dieser Flora uninteressant werden lassen.
- Das Auftreten der Zosterophyllophyten verlagert sich deutlich „nach unten“ und ist noch markanter als zuvor. Denn die von KOTYK et al. (2002) beschriebene Bathurst-Flora weist eine beachtliche Vielfalt auf und einige der dort beschriebenen Zosterophyllophyten gehören zu abgeleiteten Formen.
- Das Vorherrschen von Zosterophyllophyten paßt nicht zu bisherigen Vorstellungen, wonach deren ährige Sporangienstände als abgeleitete Merkmale gelten. Für KOTYK et al. (2002, 1012) stellt sich daher die Frage, ob dieses Merkmal als primitiv gelten muß. Dies stünde dann in direktem Widerspruch zur altehrwürdigen Telomtheorie, wonach ährige Sporangienstände durch allmähliches Zusammenrücken einzelner fertiler sog. Telome (Ursprosse) entstanden sein sollen.
- Zwar sind deutlich einfacher gebaute (aber nur sehr unvollständig überlieferte) Rhyniophytoiden nach wie vor teilweise älter als die Bathurst-Flora; dennoch tauchen mit den neuen Funden vergleichsweise komplexe Pflanzen erstaunlich früh und in beachtlicher Vielfalt auf.
- Da einige Merkmale der Bathurst-Flora wie der spezialisierte Öffnungsmechanismus der Sporangien von Macivera als fortschrittlich gelten, wären evolutionstheoretisch gesehen Vorläufer in noch tieferen Schichten zu erwarten. Für KOTYK et al. (2002) ist die Suche nach diesen Formen jedenfalls dadurch motiviert.
- Auffallend ist auch, daß diese frühen Formen teilweise Mosaikcharakter aufweisen, also Merkmale kombinieren, die für sonst eher separat in verschiedenen Gruppen vorkommen. Macivera wird als Beispiel in Abb. 1 vorgestellt. Solche Formen eignen sich allerdings nicht als Übergangsformen, da ihre Merkmale keine primitiven Vorstufen, sondern Kombinationen voll ausgeprägter Merkmale verschiedener Gruppen darstellen (vgl. dazu die Diskussion in JUNKER 1996, 69ff.). Damit erhöhen sie die Vielfalt der plötzlich auftretenden Formen und erschweren Rekonstruktionen des Evolutionsverlaufs.
Die neuen Funde sind insofern nicht überraschend, als Sporenfunde, die von Landpflanzen stammen oder stammen könnten, schon aus viel tieferen Schichten überliefert sind. Deren Zuordnung zu Makrofossilien ist allerdings oft schwierig. Man darf also auf weitere Funde gespannt sein.
|
Literatur
- BANKS HP (1975)
- Reclassification of Psilophyta. Taxon 24, 401-413.
- BANKS HP (1992)
- The classification of early land plants - revisited. The Palaeobotanist 41, 36-50.
- CAI CY, OUYANG S, WANG Y, FANG Z, RONG J, GENG L & LI X (1996)
- An early silurian vascular plant. Nature 379, 592.
- JUNKER R (1996)
- Evolution früher Landpflanzen. Studium Integrale. Neuhausen-Stuttgart.
- KENRICK P & CRANE PR (1997)
- The origin and early diversification of land plants: a cladistic study. Washington, DC.
- KOTYK ME, BASINGER JF, GENSEL PG & DE FREITAS TA (2002)
- Morphologically complex plant macrofossils from the Late Silurian of Arctic Canada. Am. J. Bot. 89, 1004-1013.
- STEWART WN & ROTHWELL GW (1993)
- Paleobotany and the evolution of plants. 2nd edition. Cambridge.
|
|
|
|  |