|
|
|
|
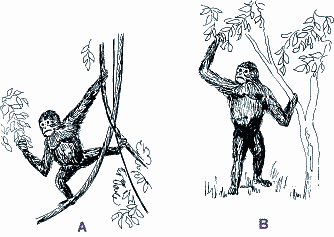
Oreopithecus bambolii ist ein ausgestorbener Affe, der in Ablagerungen des Obermiozäns der Toskana und Sardiniens (Italien) entdeckt wurde. Seine Körpergröße wird mit ca. 110-120 cm (Straus 1963) und sein Körpergewicht mit 30 kg (Fleagle 1988) bis 40 kg (Straus 1963) angegeben. Seit der Erstbeschreibung eines Unterkiefers mit Zähnen durch Gervais 1872 ist dieser fossile Affe taxonomisch umstritten. Oreopithecus wurde als naher Verwandter der cercopithecoiden Primaten (Hundsaffen) oder Hominoiden (Menschenähnlichen), als eine Form zwischen den beiden Gruppen ("forme de passage"), als Vertreter einer eigenen Hominoidenfamilie (Oreopithecidae) oder sogar als ein Hominide (Menschenartiger) eingestuft (Delson 1986, Harrison 1987). Oreopithecus besitzt einige Merkmale (kurze Hüftknochen, geringer Abstand zwischen Hüftgelenkspfanne und Darmbein-Kreuzbeingelenk, Neigung des distalen Oberschenkelknochens nach innen, prominenter vorderer unterer Darmbeinstachel), die im Zusammenhang mit einer Bipedie (Zweibeinigkeit) interpretiert wurden (Hürzeler 1949, Straus 1962, 1963, Kummer 1965). Diese Deutung konnte sich allerdings nicht durchsetzen. Die bipeden Merkmale wurden im Zusammenhang mit anderen Merkmalen (weiter Brustkorb, kurzer Rumpf, hoher Intermembranalindex) als Anpassung an vertikales Klettern gedeutet und Oreopithecus entsprechend zeichnerisch dargestellt (Abb. 1A) |
|
Kürzlich haben Köhler & Moyà-Solà (1997) zahlreiches Skelettmaterial von Oreopithecus aus dem Naturhistorischen Museum Basel untersucht. Das Forscherehepaar ist zu dem aufsehenerregenden klaren Ergebnis gekommen, daß Oreopithecus ein Affe war, der sich biped, d.h. auf zwei Beinen fortbewegte (Abb. 1B). Diese Fortbewegungsweise praktizierte Oreopithecus nicht nur gelegentlich, sondern habituell (gewohnheitsmäßig). |
Oreopithecus bambolii besitzt zahlreiche Anpassungen an eine zweibeinige Fortbewegung im Bereich der Lendenwirbelsäule, des Beckens, des Oberschenkelknochens und des Fußes (Köhler & Moyá-Solá 1997). Die Lendenwirbelsäule zeigt eine zum Bauch hin gerichtete Krümmung (Lordose). Durch diese Krümmung liegt das Zentrum der Rumpfmasse über den Kreuzbein-Darmbeingelenken und den Hüftgelenken. Dadurch ist eine nur relativ geringe Muskelanstrengung notwendig, um die Rumpfmasse beim Gehen auf zwei Beinen auszubalancieren (Latimer & Ward 1993). Die Oberschenkelknochen von Oreopithecus stehen schräg nach unten innen (X-Bein-Stellung). Die Knie befinden sich dadurch näher an der Schwerkraftlinie als bei den heute lebenden Großaffen. Das Gewicht von Oreopithecus konnte so während des zweibeinigen Gehens besser abgestützt werden. Der Fuß von Oreopithecus ist in seiner Struktur und Funktion einzigartig unter den Primaten. Üblicherweise verläuft bei den Primaten die Fußhebellinie parallel zum dritten Mittelfußknochen. Bei Oreopithecus verläuft diese Linie, bedingt durch die nach außen abgespreizten Mittelfußknochen, zwischen dem ersten und zweiten Mittelfußknochen. Zusammen mit weiteren Merkmalen an den Fußwurzelknochen (Fersenbein, Sprungbein, Würfelbein, mittleres und äußeres Keilbein) ergibt sich, daß der Fuß von Oreopithecus eine stabile Körperhaltung auf zwei Beinen ermöglichte. (Weitere Einzelheiten zu den bipeden Anpassungen von Oreopithecus siehe Kasten und Abb. 2-4.) |
LendenwirbelsäuleOreopithecus besitzt in der Lendenwirbelsäule Merkmale, von denen auf eine lumbale Lordose (Krümmung nach vorn) geschlossen werden kann. (1) Die Lendenwirbelkörper weisen eine menschenähnliche Keilform auf. (2) Die Abstände der paarigen Wirbelbogengelenke nehmen von oben nach unten zu und weisen so eine pyramidale Anordnung auf (Abb. 2). Ohne diese Anordnung würden die kleinen Wirbelbogengelenke bei lumbaler Lordose an die dazwischenliegenden Wirbelstrukturen anstoßen (Latimer & Ward 1993). Außerdem hat sie eine breite trianguläre Konfiguration an der Basis der lumbosakralen Wirbelsäule zur Folge. Diese ermöglicht eine stabile Balance des habituell aufrechten Rumpfes (Sanders 1998).
Durch die lumbale Lordose liegt die Körpermasse mit dem Zentrum über den Kreuzbein-Darmbeingelenken und Hüftgelenken und kann so beim bipeden Gehen leicht in der sagittalen Ebene ausbalanciert werden (Latimer & Ward 1993). Die Großaffen besitzen im Gegensatz zu Australopithecus und dem Menschen keine Lendenwirbelsäulenlordose. BeckenDas Schambein von Oreopithecus ähnelt in Größe und Form dem von Australopithecus afarensis. Es weist eine extrem kurze Schambeinfuge und einen geraden unteren Rand auf. Am kurzen Sitzbein von Oreopithecus befindet sich eine große Spina ischiadica (Sitzbeinstachel). Dieser Knochensporn ist beim Menschen gut, bei den Großaffen kaum und bei Australopithecus afarensis (AL 288-1) und Australopithecus africanus (Sts 14) mäßiggradig ausgebildet (Abitbol 1988, Abb. 3). Zwischen dem Ausprägungsgrad der Spina ischiadica und der Körperhaltung gibt es einen Zusammenhang. Die Bänder und Muskeln des Beckenbodens setzen direkt oder indirekt an der Spina ischiadica an. Der Beckenboden ist bei vertikaler Körperhaltung stärker belastet, da er die Eingeweide abstützt. Gut entwickelte Sitzbeinstacheln weisen somit auf eine (mehr) aufrechte Körperhaltung hin. Neben dem Ausprägungsgrad geben auch Ausrichtung und Position des Sitzbeinstachels Hinweise auf die Belastung des Beckens. Die gut entwickelten Sitzbeinstachel des Menschen sind nach innen gerichtet und befinden sich weiter vorn als bei den Großaffen. Bei den Australopithecinen sind die mäßig entwickelten Sitzbeinstachel nicht so weit vorn lokalisiert (Abitbol 1988).
OberschenkelknochenBeim Menschen ist der Oberschenkelknochen distal nach schräg innen gerichtet. Die Knie befinden sich dadurch nahe der Schwerkraftlinie und das Körpergewicht kann so während der zweibeinigen Fortbewegung gut abgestützt werden. Der Oberschenkelknochen der Pongiden ist dagegen distal nur gering nach innen oder sogar leicht nach außen geneigt. Dadurch ist es ihnen möglich, den Greiffuß unterhalb des Körperschwerpunktes zu plazieren, während das Knie stark abgespreizt ist (Preuschoft & Tardieu 1996). Der Oberschenkelknochen von Oreopithecus ähnelt in diesem Merkmal mehr dem Oberschenkelknochen des Menschen und dem von Australopithecus (Köhler & Moyà-Solà 1997). Es kann deshalb vermutet werden, daß die Kniegelenke von Oreopithecus näher an der Schwerkraftlinie und damit günstiger für eine zweibeinige Fortbewegung positioniert waren, als dies bei den heute lebenden Großaffen der Fall ist. Vom distalen Neigungsgrad des Oberschenkelknochens kann allerdings nicht unmittelbar auf die genaue Position des Kniegelenkes in Bezug auf die Schwerkraftlinie geschlossen werden. Eine Reduktion der Oberschenkellänge oder eine Vergrößerung des Abstandes zwischen den beiden großen Rollhügeln der Oberschenkelknochen (größerer Abstand zwischen den Hüftgelenkspfannen, spitzerer Winkel zwischen dem Hals und dem Schaft des Oberschenkelknochens, längerer Oberschenkelhals) erfordern für die gleiche Position der Kniegelenke eine stärkere X-Beinstellung gegenüber dem Menschen (Preuschoft & Tardieu 1996). Alle vier genannten Merkmale sind bei den Australopithecinen, wenn auch in unterschiedlichem Maße, gegeben. Es ist deshalb sehr fraglich, ob die Kniegelenke dieser frühen Hominiden trotz ihrer Hyper-X-Beinstellung (distale Oberschenkelschaftneigung 2-3 Grad höher als beim Menschen; Heiple & Lovejoy 1971, Tardieu & Trinkaus 1994) eine menschliche Position einnahmen. Ein Rückschluß von der distalen Oberschenkelneigung auf die genaue Art der bipeden Fortbewegung ist nicht möglich (Jenkins 1972, siehe auch Brandt 1995). FußDer Fuß von Oreopithecus weist ein unter den Primaten einzigartiges Merkmalsmosaik auf (Abb. 4).
Am auffälligsten an diesem Fuß ist die nach innen verlaufende Fußhebellinie. Üblicherweise verläuft die Fußhebellinie bei den Primaten parallel zum dritten Mittelfußknochen. Bei Oreopithecus verläuft sie dagegen zwischen dem ersten und zweiten Mittelfußknochen wie beim Menschen. Im Gegensatz zum Menschen wird dieser Fußhebellinienverlauf aber durch abduzierte (abgespreizte) äußere Mittelfußknochen hervorgerufen (Abb. 4). Wenn ein Gorilla sich auf dem Erdboden bewegt, dann sind seine Mittelfußknochen ähnlich nach außen abgespreizt. Auch die geringe Größe des Würfelbeines, die großen Gelenkflächen auf dem äußeren und mittleren Keilbein und der zwischen den Keilbeinen und dem dritten Mittelfußknochen verankerte zweite Mittelfußknochen weisen auf eine hohe Kraftübertragung im Innenbereich des Fußes hin. Die Mittelfußknochen sind kurz, nur gering gekrümmt, und ihre Länge nimmt nach innen zu ab. Zusammen mit dem nach innen verlaufenden Fußhebel hat das zur Folge, daß während der Fortbewegung auf zwei Beinen nur geringe Scherkräfte im Mittelfußbereich auftreten. Außerdem ist damit das Abstützgebiet des Fußes für die Ausbalancierung des Körperschwerpunktes beim zweibeinigen Stehen und Gehen sehr groß. Die Ausrichtung des Fersenbeinhöckers vertikal zum Untergrund und in einem nahezu rechten Winkel zur oberen talaren Oberfläche sowie die ähnliche Höhe des inneren und äußeren Randes des Sprungbeines sind ebenfalls bipede Merkmale. Sie lassen darauf schließen, daß die Schienbeinachse in einer Linie mit der Schwerkraftlinie ausgerichtet war. Auch die Fußproportion mit einem hohen Kraftarm/Lastarmverhältnis und einige Gelenkmerkmale im Fußwurzel-Mittelfußbereich von Oreopithecus sprechen für eine bipede Fortbewegung und gegen Kletteraktivitäten bei diesem Affen. |
Sardinien und die Toskana waren im Obermiozän eine Insel in einem Ozean, der sich von Südeuropa bis Südostasien erstreckte. Das heutige Mittelmeer ist der Rest dieses Meeresbandes. Oreopithecus ist somit ein endemischer Primat einer mediterranen Insel. Solche Inseln weisen zwei Charakteristika auf: Es fehlen Räuber und es existiert eine Begrenzung des Raumes und der Nahrungsressourcen. Angesichts des Fehlens von Räubern waren bei Oreopithecus Fortbewegungsanpassungen an das energetisch aufwendige und risikovolle Klettern in Bäumen, um sich diesen entziehen zu können, nicht notwendig (Köhler & Moyà-Solà 1997). Der eigentümliche Fuß und die kurzen Beine von Oreopithecus sind für schnelles Gehen oder Laufen wenig geeignet. Jedoch war eine stabile Körperhaltung auf zwei Beinen möglich. Bipedes Stehen beim Greifen nach Nahrung kombiniert mit häufigem zweibeinigem Watscheln über kurze Strecken bei der Nahrungssuche, wurde von Hunt (1994) auch bei Wildschimpansen beobachtet. Dies könnte auch bei Oreopithecus eine sehr effektive Strategie zur Nahrungsgewinnung gewesen sein (Köhler & Moyà-Solà 1997). Oreopithecus bambolii lebte in sumpfigen Waldgebieten (Delson 1988). Meistens wird er als Blattfresser beschrieben (Szalay & Delson 1979, Delson 1988, Fleagle 1988). In einer neueren Veröffentlichung vemuten Moyà Solà & Köhler (1997) jedoch, daß Oreopithecus eher ein Allesfresser war. |
Der Bau des Körperstamm- und Extremitätenskeletts von Oreopithecus bambolii zeigt deutliche Anpassungen an eine zweibeinige Fortbewegung auf dem Erdboden. Oreopithecus ist aus mehreren Gründen als Hominide wenig geeignet. Der letzte gemeinsame Vorfahre des Schimpansen und des Menschen wird derzeit vor 6 Millionen Jahren oder etwas früher vermutet. Oreopithecus besaß ein Alter von 7-9 Millionen radiometrischen Jahren. Das mediterrane Lebensgebiet von Oreopithecus war geographisch isoliert und weit entfernt von Afrika, dem Verbreitungsgebiet der allgemein anerkannt frühesten Hominiden, den Australopithecinen. Außerdem sprechen auch einige spezielle morphologische Anpassungen gegen eine stammesgeschichtliche Beziehung zum Menschen. Einige bipede Anpassungen von Oreopithecus wie die lumbale Lordose, gut ausgebildete Sitzbeinstachel und die schräge untere Femurstellung werden im derzeitigen evolutionstheoretischen Rahmen als Konvergenzen mit den Australopithecinen und Menschen gedeutet. Oreopithecus war ein ganz speziell angepaßter miozäner Menschenaffe. Das Beispiel Oreopithecus lehrt eindrucksvoll, daß Anpassungen an eine Fortbewegung auf zwei Beinen auf dem Erdboden, auch bei Menschenaffen, nicht zwangsläufig eine Vorfahrenschaft zum Menschen begründen. Die Australopithecinen werden heute von den Evolutionstheoretikern in die Vorfahrenschaft des Menschen gestellt. Ein wesentlicher Grund für diese Deutung sind anatomische Anpassungen an eine Fortbewegung auf zwei Beinen. Diese Tiere haben jedoch eine ganz eigene Art der zweibeinigen Fortbewegung praktiziert, von der der menschlich-schreitende Gang nicht ohne weiteres abgeleitet werden kann (Brandt 1995). Die zweibeinige Fortbewegung des miozänen Menschenaffen Oreopithecus bambolii zeigt, daß eine bipede Fortbewegung mehrfach unabhängig in verschiedenen Gruppen aufgetreten ist. Dieser Befund kann als Bestätigung für die Deutung der Australopithecinen als eine eigenständige Gruppe von Großaffen ohne historisch-verwandtschaftliche Beziehung zum Menschen gewertet werden. |

|