 |
 Studium Integrale Journal
Studium Integrale Journal
21. Jahrgang / Heft 2 - Oktober 2014
Seite 116 - 123
|
|
|
Der Fund eines fossilen Königsfarns aus dem schwedischen Unterjura (180 Millionen radiometrische Jahre) stellt bisher bekannte Beispiele lebender Fossilien aufgrund detaillierter Erhaltung selbst von subzellulären Strukturen in den Schatten. Dass auch zelluläre Details über einen mutmaßlich so großen Zeitraum nahezu unverändert bleiben, wirft einige Fragen auf.
Lebewesen, die als Fossilien bekannt sind, aber auch heute in sehr ähnlicher Form existieren, üben eine besondere Faszination aus. Der Zahn der Zeit scheint ihnen nichts anhaben zu können. Solche „lebenden Fossilien“ werden in evolutionärer Perspektive eher als Ausnahmen angesehen – widerstanden sie doch viele Millionen Jahre lang dem scheinbar unausweichlichen Drang der Natur zur Veränderung, der – so die Ansicht der Evolutionsbiologie – zur immensen Vielfalt an Formen geführt haben soll. Wenn also Evolution das Normale ist, wären unverändert lebende Fossilien als Ausnahme zu kennzeichnen. Was aber, wenn die Ausnahme zur Regel wird? Denn lebende Fossilien sind keineswegs so selten, mindestens wenn man den Vergleich zwischen heute lebend und fossil nicht zu eng auf der Artebene ansetzt, sondern auf der Ebene von Grundtypen. Wie kommt es, dass der mutmaßlich lang anhaltende Prozess der Evolution bei so vielen Organismen irgendwann zum weitgehenden Dauerstillstand gekommen ist? Ist eine innovative Evolution wirklich eine Grundeigenschaft des Lebens? Oder bestätigen lebende Fossilien nicht gerade das Gegenteil, dass den Veränderungen der Organismen Grenzen gesetzt sind? Aus der Sicht der Schöpfungslehre ist das häufige Vorkommen lebender Fossilien nicht überraschend, ja dies kann sogar als eine Voraussage verstanden werden, die durch Forschung geprüft werden kann.
Auch wenn lebende Fossilen also keineswegs selten sind, ist der Fund eines „lebenden Fossils“ eines Königfarns, über den Bomfleur et al. (2014) berichten, bisher einzigartig und höchst erstaunlich. Dass Königsfarne (Familie Osmundaceae) sich seit dem unteren Mesozoikum (220 Millionen radiometrische Jahre) in ihrem Bau fast nicht verändert haben, ist schon länger bekannt. Der jüngst beschriebene Fund aus dem schwedischen Unterjura (180 Millionen radiometrische Jahre) stellt bisher bekannte Beispiele lebender Fossilien aufgrund detaillierter Erhaltung selbst von subzellulären Strukturen jedoch in den Schatten. Ein etwa 6 cm langes Rhizom (Erdspross) von ca. 7 mm Durchmesser und zahlreichen Ansätzen von Blattwedeln lässt aufgrund besonderer Erhaltung nicht nur die Zellwände sehr gut erkennen, sondern auch Zellkerne und Zellorganellen. Und es kommt noch besser: Einige der Zellen waren gerade in Teilung begriffen, so dass sogar spezifische Chromosomen in verschiedenen Teilungsstadien identifiziert werden können (Abb. 1). Die Größe der fossilen Zellkerne während der Interphase entspricht der Größe heutiger Osmundaceae. Die Autoren schließen daraus, dass sich die Genomgröße 180 Millionen Jahre lang nicht geändert hat, ein „hervorragendes Beispiel eines evolutionären Stillstands“ (Bomfleur et al. 2014, 1376). Weder Vervielfachung des Erbguts noch nennenswerte Genverluste seien vorgekommen – der evolutionäre Stillstand ist nahezu total. „Wenn wir die feinen Zellstrukturen dieses 180 Millionen Jahre alten Fossils mit denen heute lebender Königsfarne vergleichen, sind sie eigentlich identisch“, wird Bomfleur im Deutschlandfunk zitiert (http://tinyurl.com/paowga7).
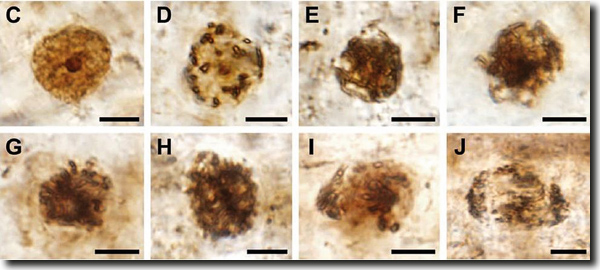 |
| Abb. 1: Zellteilungsphasen im beschriebenen fossilen Königsfarn. C Kern der Interphase mit Nucleolus und intakter Kernmembran, D Früher Prophase-Kern mit kondensierendem Chromatin und sich auflösendem Kern und Kernmembarn, E und F späte Prophase-Zellen mit aufgewickelten Chromosomen, Nucleolus und Kernmembran vollständig aufgelöst, G and H Prometaphase-Zellen; Chromosomen ordnen sich am Kern-Äquator an, I und J mögliche Anaphase-Zellen, Chromosomen sind in Richtung der Pole. (Aus Bomfleur et al. 2014) |
|
Möglich wurde die hervorragende Erhaltung dadurch, dass durch eine vulkanische Schlammlawine vermutlich 70-120˚C Grad Celsius heiße, mit Mineralen beladene vulkanische Wässer Kontakt mit den Pflanzen bekamen und dadurch Kalzit auskristallisierte. Das ermöglichte die Konservierung des Inhalts der Zellen bis in feinste Details. Die Fossilisierung muss ungewöhnlich schnell innerhalb weniger Minuten abgelaufen sein.
Lebende Fossilien wie die Königsfarne sind eine Herausforderung für den Geltungsanspruch einer evolutionsbiologischen Interpretation des Lebens. Darüber hinaus ergeben sich aber auch Anfragen dahingehend, ob die ermittelten Zeiträume wirklich reale Zeitabschnitte repräsentieren. Diese Anfrage wiegt umso schwerer, je vehementer man auf die Evolution als eine Grundeigenschaft des Lebens verweist.
[Bomfleur B, McLoughlin S & Vajda V (2014) Fossilized nuclei and chromosomes reveal 180 million years of genomic stasis in royal ferns. Science 343, 1376-1377.]
|

|
Mehrfach wurde in Studium Integrale Journal schon über unerwartet moderne Eigenschaften kambrischer Lebewesen berichtet. Zuletzt wurden komplexe Nervensysteme bei den Gliederfüßern Alacomenaeus und Fuxianhuia protensa dokumentiert. Nun sorgt ein weiteres Fuxianhuia-Fossil für Furore.
Nachdem bei dieser auf 520 Millionen radiometrische Jahre datierten Art schon der Verdauungstrakt und das komplexe Nervensystem mit modern anmutendem dreigliedrigem Gehirn versteinert aufgefunden worden war (Ma et al. 2012), konnte nun an einem wirklich außergewöhnlichen Fossil das bisher älteste komplett konservierte Blutgefäßsystem im Detail untersucht werden. Das Fossil „YKLP 11335“ stammt aus einer Tonsteinschicht, in der normalerweise nur schlecht konservierte Organismen gefunden werden. Generell ist das Exemplar auch nicht gut erhalten, allein an einer Art hellem Negativ-Umriss ist die segmentierte Form der Art Fuxianhuia protensa identifizierbar. Seltsamerweise ist ausgerechnet und fast ausschließlich das fragile Blutgefäßsystem deutlich als schwarze fein gegliederte Struktur auf dem hellen Hintergrund erhalten geblieben, vergleichbar fast einer Detailzeichnung im Lehrbuch. In der Mitte ist das längliche Herz erkennbar, von dem analog zur äußerlichen Gliederung in Segmente regelmäßig Blutgefäße zu den Seiten abgehen. Gut erkennbar sind auch kräftige Arterien zur Blutversorgung des Gehirns, der Augen und der Antennen. Diese effiziente Versorgung von Sinnesorganen und Signalverarbeitung bringt die Autoren Ma, Edgecombe und Strausfeld (2014) dazu, Fuxianhuia protensa als sehr beweglichen kambrischen Jäger mit gutem Seh- und Geruchssinn zu interpretieren. Interessant ist, dass die Autoren die fragliche fossile Struktur explizit durch Vergleich mit heute lebenden Gliederfüßern klar als Blutgefäßsystem identifizieren, denn seine Anordnung entspricht der, die man in vielen heutigen terrestrischen und aquatischen Gliederfüßern findet.
 |
| Abb. 1: Fuxianhuia protensa in dorsaler (rückenseitiger) Sicht. (Aus Ma et al. 2014) |
|
Fuxianhuia protensa sieht äußerlich vielleicht primitiv aus, ihre inneren Qualitäten weisen die Art nun in mehrfacher Hinsicht als modern aus, was die Autoren bei der Diskussion der angenommenen Evolution der Gliederfüßer vor Schwierigkeiten stellt: Ist die Art, wie man von ihrem Alter und Äußeren her annehmen könnte, eine ursprüngliche Stammform, müssten heutige Gliederfüßer-Arten mit niedrigerem Komplexitätsgrad, die es durchaus auch gibt, effiziente Strukturen wieder verloren haben. Wäre Fuxianhuia dagegen wegen seines komplexen Inneren nicht Vorläufer der heutigen Gliederfüßer, müssten letztere ihre komplexe Organisation unabhängig noch einmal entwickelt haben. Beides sind Beispiele für unplausible Szenarien, zu denen Evolutionsbiologen oft durch den Datenbefund gezwungen werden, wenn eine gemeinsame Abstammung von einfachen Stammformen vorausgesetzt wird. Dieses kambrische Fossil entspricht einmal mehr offensichtlich nicht dem theoretisch erwarteten Prinzip der Evolution von „einfach“ zu „komplex“. Im Gegenteil, ganz am Beginn ihres Erscheinens im Fossilbericht im frühen Kambrium, zu Zeiten der berühmten „kambrischen Explosion“, besaßen Gliederfüßer schon höchst komplex entwickelte Nerven- und Blutgefäßsysteme, die sich dann über eine halbe Milliarde radiometrische Jahre offensichtlich nicht mehr geändert haben. Es kann auch hier wieder konstatiert werden, dass ein solcher Mix aus „ursprünglichen“ und „modernen“ Strukturen innerhalb einer Art einfacher schöpfungstheoretisch, im Sinne eines der geschaffenen Natur zugrunde liegenden „Baukastensystems“ gedeutet werden kann, dessen Komponenten keine größeren Abwandlungen über die Zeit mehr erfahren.
[Ma X, Peiyun C, Hou X, Edgecombe GD & Strausfeld NJ (2014) An exceptionally preserved arthropod cardiovascular system from the early Cambrian. Nature Communications 5:3560; doi: 10.1038/ncomms4560; Ma X, Hou X, Edgecombe GD&Strausfeld NJ (2012) Complex brain and optic lobes in an early Cambrian arthropod. Nature 490, 258-261, doi:10.1038/nature11495, Harvey THP, Vélez MI & Butterfield NJ (2012) Exceptionally preserved crustaceans from western Canada reveal a cryptic Cambrian radiation. Proc. Natl. Acad. Sci. 109, 1589-1594, doi: 10.1073/pnas.1115244109]
|
|
|
Die kambrischen Gesteinsschichten bergen eine große Anzahl verschiedenster Baupläne, die recht abrupt in der Fossilüberlieferung erscheinen. Die dort auch vertretenen Anomalocariden galten bisher als furchterregende Räuber. Nun hat sich herausgestellt, dass einige von ihnen Filtrierer waren, die Kleinstlebewesen aus dem Wasser siebten. Damit sind sie ein weiterer Baustein der kambrischen Tier-Vielfalt und wieder einmal ein Beispiel für Konvergenz – die mehrmalige unabhängige Entstehung eines komplexen Bauplanelements.
Im Jahr 1989 beschrieb der unvergessene Paläontologe Stephen J. Gould in seinem Buch „Wonderful Life“ (deutsch 1991) die ungewöhnliche Tierwelt des Kambriums, besonders des mittelkambrischen Burgess-Schiefers in Britisch-Kolumbien (Kanada). Unter dessen Fossilien klassifizierte er 20 neue Tierstämme mit großen Bauplan-Unterschieden, die er als „irre Wundertiere“ bezeichnete; die äußerst vielgestaltigen Tiere nannte er „einzigartige Gliederfüßer“ mit einem „Maximum an anatomisch leistungsfähigen Möglichkeiten“. Seiner Meinung nach übertreffen die Burgess-Fossilien „wahrscheinlich das gesamte Spektrum des wirbellosen Lebens in den heutigen Ozeanen“.
 |
| Abb. 1: Vordere Körperanhänge von Tamisiocaris borealis von der Sirius Passet-Lagerstätte (Unterkambrium Nordgrönland). a Isolierter Körperanhang, b Isolierter Körperanhang mit den ansitzenden Stacheln im Detail, c Details der Stacheln (vergrößert aus b). (Aus Vinther et al. 2014) |
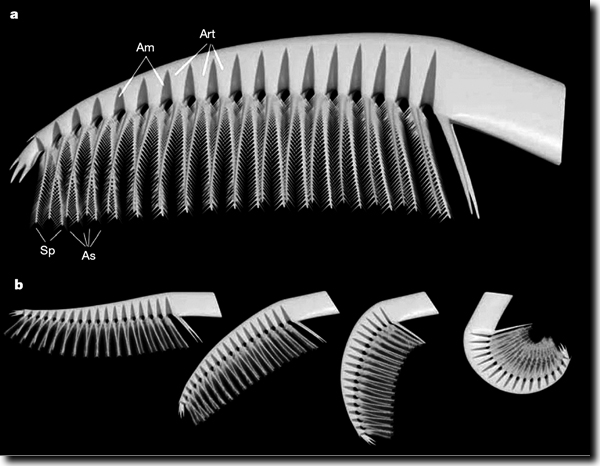 |
| Abb. 2: Rekonstruktion von Anhängen von Tamisiocaris. a Einzelner Anhang mit Membranen (Am), einzelnen Abschnitten (Art), Stacheln (Sp) und Hilfsstacheln (As). b Mutmaßliche Abfolge der Bewegungen der Vorderen Anhänge von Tamisiocaris. (Aus Vinther et al. 2014) |
|
Zu diesen „irren Wundertieren“ gehören die Anomalocariden („ungewöhnliche Garnelen“). Mit einer Länge von bis zu zwei Metern gehörten sie zu den größten Tieren der kambrischen Gewässer. Sie werden zu den Gliederfüßern (Arthropoda) gerechnet. Mit ihren Seitenlappen am segmentierten Rumpf waren sie wahrscheinlich gute und wendige Schwimmer.
Die Anomalocariden galten bislang als formidable Räuber, doch neuere Untersuchungen stellen das teilweise in Frage und liefern ein überraschendes Ergebnis. Neue Fossilfunde aus Nordgrönland legen nahe, dass einige Arten der Anomalocariden sich mit Hilfe einer ausgeklügelten Filtriertechnik von Plankton ernährten – ähnlich wie die heutigen Bartenwale wie z. B. der Blauwal. Untersucht wurden fünf fossil erhaltene, stachelbewehrte, ca. 12 cm lange Anhänge und zwei mit dem Kopfschild verbundene Anhänge von Tamisiocaris borealis aus der unterkambrischen Sirius-Passet-Konservat-Lagerstätte. Dabei wurden die beiden langen, stachelbewehrten Anhänge am Kopf genauer unter die Lupe genommen. Die Greifer erwiesen sich als beweglich und die filigranen Fortsätze besitzen feine, ineinander greifende Stacheln. Diese interpretieren die Forscher als Werkzeuge, mit deren Hilfe Tamisiocaris Organismen ab einer Größe von etwa 1/2 Millimeter aus dem Wasser gefischt hat (Vinther et al. 2014). „Ausgestreckt bilden sie eine Art Kamm, den der Urzeit-Gliederfüßer durch das Wasser gezogen haben könnte wie eine Art Netz. Rollte er dann die Anhänge zum Mund hin ein, bildeten ihre Borsten einen dichten Käfig, in dem gefangene Organismen hängenblieben“ (Podbregar 2014).
Aufgrund der Filtrierer-Fähigkeit stellen die Forscher Tamisiocaris in die neue Gruppe der Cetiocaridae (Walgarnelen; cetus, lat. = Wal; „caris“ steht für Garnele). Die Autoren schließen, dass ein vielfältiges Ökosystem mit gut entwickelten Nahrungsketten in den offenen Meeren ausgeprägt gewesen sei. Denn um als Filtrierer leben zu können, ist eine hohe Primärproduktion und ausreichend Plankton Voraussetzung. Vinther et al. (2014) geben auch eine Übersicht über die Vielfalt des Baus der Anhänge der Anomalocariden, z. B. solche mit dreizackigen Stacheln oder mit scherenartigen Anhängen und stellen fest (S. 498), dass die außerordentliche morphologische Vielfalt der Anhänge zeige, dass es sich bei den Anomalocariden nicht um ein gescheitertes Evolutionsexperiment handle, sondern um eine umfangreiche Radiation im Laufe der kambrischen Explosion.
Die Untersuchung ist in zweierlei Hinsicht bemerkenswert. Zum einen vergrößern die Beobachtungen an Tamisiocaris die Vielfalt an ausgeklügelten Einrichtungen, die bereits im Unterkambrium ohne erkennbare Vorfahren präsent waren. Zum anderen zeigen sie einmal mehr, dass komplexe Einrichtungen mehrfach (oft vielfach) unabhängig auftauchen – in diesem Fall die Fähigkeit der Nahrungsfiltrierung. Eine ganze Reihe von Filtrierern gibt es außer den bereits erwähnten Bartenwalen auch unter den Gliederfüßern; und die Filtrierer sind nun um ein interessantes Mitglied erweitert worden, das diese Fähigkeit unabhängig erworben haben muss (Konvergenz). Vinther et al. (2014) erwähnen, dass bei den Cetiocaridae ähnliche Einrichtungen zum Filtrieren verwirklicht sind wie bei anderen Filtrierern unter den Gliederfüßern (z. B. den Rankenfußkrebsen, Cirripedia). Dabei muss bedacht werden, dass diese Fähigkeit sehr anspruchsvoll ist und viele Details zusammenstimmen müssen. Ein allmählicher Erwerb oder – wie man evolutionstheoretisch vermutet – ein Umbau von einer räuberisch lebenden Ausgangsform ist sehr fragwürdig. Denn um ernährungsmäßig über die Runden kommen zu können, muss die Filtrierfunktion gut ausgebaut sein. Und der Verlust des räuberischen Nahrungserwerbs kann selektiv wohl kaum begünstigt werden, solange die neue Art des Nahrungsgewinns nicht ausreichend funktioniert.
Vinther et al. (2014, 499) dagegen werten die Existenz der filtrierenden Gattung Tamisiocaris und das Ausmaß unabhängiger (konvergenter) Entstehung ähnlicher Konstruktionen als Beleg für Vorhersagbarkeit und Kanalisierung der Evolution. Worin diese Kanalisierung aber bestehen soll, ist völlig unklar und wird nicht thematisiert.
Intelligente Strukturen können aus wissenschaftlicher Perspektive als Hinweis auf eine intelligente Verursachung gewertet werden. Auch wenn man über den Prozess der Designentstehung naturwissenschaftlich nichts aussagen kann, bedeutet dies nicht gleichzeitig, dass der Schluss auf einen übernatürlichen Entstehungsprozess deshalb unberechtigt wäre.
[Gould SJ (1989) Wonderful Life. New York: W. W. Norton & Co. (dt. 1991: Zufall Mensch. München – Wien); Vinther J, Stein M, Longrich NR & Harper DAT (2014) A suspension-feeding anomalocarid from the Early Cambrian. Nature 507, 496-500; Podbregar N (2014) Sanfte Riesen im Urzeitmeer. http://www.wissenschaft.de/erde-weltall/palaeontologie/-/journal_content/56/12054/3250966/Sanfte-Riesen-im-Urzeitmeer/]
|
|
Belgica antarctica ist der wissenschaftliche Name einer Zuckmücke (Familie Chironomidae), die als einziges Insekt beschrieben ist, dessen Lebensraum ausschließlich die Antarktis ist (endemische Verbreitung). In diesem Ökosystem ist sie extremen Lebensbedingungen ausgesetzt, wie z. B. tiefen Temperaturen, wiederkehrender ausgeprägter Trockenheit, hoher UV-Strahlung etc. Die Larven, die während ihrer zweijährigen Entwicklung die meiste Zeit des Jahres in Eis eingeschlossen sind, entwickeln sich zu Beginn ihres dritten Sommers zu flügellosen erwachsenen Mücken. Diese suchen krabbelnd Fortpflanzungspartner, paaren sich, legen Eier und sterben nach 7-10 Tagen.
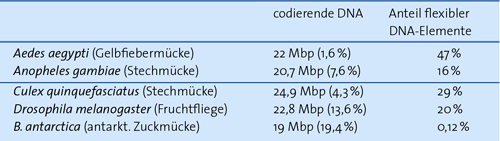 |
| Tab. 1: Absolute Größe der codierenden DNA und ihr relativer Anteil am Gesamtgenom sowie die Anteile flexibler DNA-Elemente bei einigen Mückenarten. (Nach Kelley et al. 2014) |
|
David L. Denlinger, unter dessen Leitung bereits eine Reihe von Studien über B. antarctica veröffentlicht worden ist, hat nun eine Untersuchung über deren Erbgut vorgelegt (Kelley et al. 2014). Das Genom ist mit 99 Millionen Basenpaaren (99 Mbp) auffällig klein, selbst das der parasitischen Kleiderlaus (Pediculus humanus humanus) ist mit ca. 105 Mbp umfangreicher, wie auch das der Fruchtfliege (Drosophila melanogaster) mit ca. 140 Mbp. Dabei codieren knapp 19 Mbp (ca. 19,4 %) für mehr als 13.500 Proteine. Die Autoren zeigen durch einen Vergleich mit anderen Mücken, dass im Genom von B. antarctica mit 19 Mbp ein hoher Anteil codierender DNA enthalten ist (Tab. 1).
Der absolute Umfang der codierenden DNA ist bei stark unterschiedlichen Genomgrößen durchaus vergleichbar, in allen Mücken liegen also ähnlich viele Gene vor. Die Kleinheit des Genoms von B. antarctica resultiert nach den vorliegenden Daten vor allem aus dem geringen Anteil an im Genom beweglichen Sequenzabschnitten (transposable elements), aus der geringen Menge sich wiederholender Sequenzbereiche und aus kürzeren und weniger in den Genen enthaltenen Einschüben (Introns).
Das von Kelley et al. (2014) präsentierte Erbgut ist das kleineste bisher bekannte Insektengenom. Die extremen Lebensbedingungen in der Antarktis stellen B. antarctica einerseits vor enorme Herausforderungen und bauen andererseits einen großen Selektionsdruck auf, der das kleine Genom erzwingt. Die antarktische Zuckmücke stellt damit ein Beispiel für die enorme Flexibilität von Lebewesen dar und ihr Erbgut könnte sich als gutes Studienobjekt für ein besseres Verständnis der nicht-codierenden DNA-Abschnitte erweisen.
[Kelley JL, Peyton JT, Fiston-Lavier A-S, Teets NM, Yee M-C, Johnston JS, Bustamante CD, Lee RE & Denlinger DL (2014) Compact genome of the antarctic midge is likely an adaption to an extreme environment. Nat. Commun. 5:4611 doi: 10.1038/ncomms5611]
|
 |
| Abb. 1: Eine heutige Augenfliege mit den charakteristisch großen Facettenaugen. (© Nikola Rahmé, Senckenberg-Gesellschaft für Naturforschung) |
 |
| Abb. 2: Bisher größte bekannte fossile Augenfliege Metanephrocerus belgardeae. (© S. Bruce Archibald, Senckenberg-Gesellschaft für Naturforschung) |
|
Augenfliegen (Pipunctulidae) sind durch ihre auffällig großen, fast den gesamt Kopf einnehmenden Augen charakterisiert. Die Weibchen legen nach dem Paarungsflug ihre Eier als Parasiten (Endoparasiten) in Wirten, meist Zikaden ab. Etwas mehr als 1400 Arten sind beschrieben (etwa 110 davon kommen in Deutschland vor) und weitere ca. 1300 werden – noch nicht wissenschaftlich erfasst – vermutet. Über die Vergangenheit der Augenfliegen weiß man aus der Fossilüberlieferung vergleichsweise wenig. Bisher sind 12 Einschlüsse in Bernstein und 5 Fossilien als Abdrücke (compression fossils) dokumentiert.
Aufgrund genetischer Studien wird das Alter für den Beginn der Entfaltung dieser Familie der Zweiflügler (Diptera) mit ungefähr 70 Millionen Jahren angegeben (Wiegemann et al. 2011). Christian Kehlmaier vom Senckenberg Museum in Dresden hat nun gemeinsam mit Kollegen in zwei Veröffentlichungen drei fossile Augenfliegen beschreiben. Zwei als Einschlüsse in Baltischem Bernstein erhaltene Formen konnten sie mit computertomographischen Methoden (Micro-CT) sehr genau untersuchen und beschreiben. In einer Pressemitteilung der Senckenberg-Gesellschaft für Naturforschung wird Kehlmaier mit folgenden Worten zitiert: „Das sind schon ganz besondere Details, die wir an den etwa 40 bis 50 Millionen Jahre alten Tieren zeigen konnten“, um dann fortzufahren: „Die Lebensweise und das Aussehen der Augenfliegen haben sich vor 70 Millionen Jahren entwickelt. Die nun neu beschriebenen fossilen Arten lassen vermuten, dass schon relativ kurz nach der Etablierung der großäugigen Tiere ein reichhaltiges Artenspektrum existiert haben muss, das dem heutigen in keiner Weise nachsteht.“ Befunde dieser Art werden vor allem bei Inklusen in fossilem Harz regelmäßig angetroffen.
Bei der Interpretation der Fossilfunde im evolutionstheoretischen Deutungsrahmen muss häufig ein schnelles Auftreten angenommen werden, dem nur noch vergleichsweise geringe Änderungen folgten.
[Archibald SB, Kehlmaier C & Mathewes RW (2014) Early Eocene big headed flies (Diptera) from the Okanagan Highlands, Western North America. Can. Entomol., doi: 10.4039/tce.2013.79; Kehlmaier C, Dierick M & Skevington JH (2014) Micro-CT studies of amber inclusions reveal internal genitalic features of big-headed flies, enabling a systematic placement of Metanephrocerus Aczél 1948 (Insecta: Diptera: Pipunculidae). Arthropod Systematics & Phylogeny 72, 23-36; Wiegmann BM, Trautwein MD et al. (2011) Episodic radiation in the fly tree of life. Proc. Natl. Acad. Sci. 108, 5690-5695.]
|
|
Die molekularen Bestandteile, die dem programmierten Zellabbau (oder Zelltod, Apoptose) zugrunde liegen, sind zwischen Mensch und Koralle austauschbar. Aus evolutionsbiologischer Perspektive muss ein erstaunliches Ausmaß an evolutionärem Stillstand (Stasis; Konservierung) postuliert werden. Ausgerechnet unter den basalen und damit als stammesgeschichtlich sehr alt geltenden Vielzellern findet sich das vielseitigste Apoptose-System.
Ein wichtiger formbildender Prozess in der Ontogenese (individuelle Entwicklung von der befruchteten Eizelle bis zum ausgewachsenen Organismus) ist der programmierte Zellabbau, also das physiologische (nicht krankhafte), programmierte Absterben von Zellen (Apoptose). Während es z. B. beim krankhaft bedingten Zelltod in der Regel zu einem unkontrollierten Aufquellen und Zerplatzen der Zelle oder zu einer unkoordinierten Zerstörung von Eiweißen oder Zellorganellen kommt (Nekrose), geschieht die Auflösung der Zellbestandteile und ihr Abbau im Rahmen der Apoptose hochgradig organisiert und physiologisch gesteuert. Apoptose dient damit immer dem Erhalt oder der koordinierten Entwicklung des Organismus. Beispielsweise werden Gewebestrukturen zwischen den ontogenetischen Anlagen von Fingern und Zehen kontrolliert zum Abbau gebracht oder durch die Auflösung der Zellen von Glaskörper und Linse wird die Lichtdurchlässigkeit der Augenlinse ermöglicht. Aber auch im ausgewachsenen Organismus spielt Apoptose eine wichtige Rolle, z. B. in der Kontrolle der Zellzahl und der Größe von Geweben, bei der Entfernung entarteter Zellen oder bei der Rückbildung der Gebärmutter nach der Geburt. Bei der Apoptose werden Nachbarzellen bzw. -gewebe durch austretende Gifte wie im Fall des o.g. krankhaft bedingten Zelltodes nicht geschädigt.
Es ist schon länger bekannt, dass die molekulare Basis der Apoptose hochkonserviert ist, das heißt, sie ist bei sehr verschiedenen Organismengruppen sehr ähnlich, was evolutionstheoretisch so interpretiert wird, dass dieser Mechanismus stammesgeschichtlich sehr früh entstand und anschließend nicht mehr nennenswert verändert wurde. In einer neuen Untersuchung wiesen amerikanische Wissenschaftler (Quistada et al. 2014) nach, dass Apoptose auch bei riffbildenden Korallen vorkommt. Diese gelten im Rahmen der Evolutionsanschauung als Repräsentanten einer der ältesten Tiergruppen. Es zeigte sich, dass alle wesentlichen Komponenten des Apoptose-Programms bei Korallen vorhanden sind und denen des Menschen gleichen. Dabei spielen die Tumornekrosefaktor (TNF)-Superfamilien (TNFRSF/TNFSF) eine zentrale Rolle. Die Wissenschaftler untersuchten 53 Proteine der TNFRSF- und TNFSF-Familien in der riffbildenden Koralle Acropora digitifera und stellten große Sequenzähnlichkeiten mit den entsprechenden Proteinen des Menschen fest, besonders bezüglich der aktiven Zentren dieser Eiweiße. Ein Vergleich mit der Taufliege Drosophila ergab, dass in dieser Linie im Gegensatz zur Linie zum Menschen wichtige Teile der TNF-Superfamilie fehlen, was evolutionstheoretisch als Verlustentwicklung interpretiert wird. Zuvor war angenommen worden, dass die Zahl der Mitglieder der TNF-Superfamilie nach der Trennung der Linie der Wirbeltiere von der der Wirbellosen zugenommen habe. Nun hat sich herausgestellt, dass im Gegenteil die Korallen das vielseitigste Repertoire der TNF-Superfamilien besitzen.
Die große Ähnlichkeit der molekularen Komponenten des Apoptose-Prozesses bei Korallen und Menschen wurde eindrucksvoll demonstriert durch die Einführung eines menschlichen TNF (HuTNF) in die Korallen. Es zeigte sich, dass es direkt an die Korallen-Zellen bindet, dadurch die Caspase-Aktivität erhöht, was (über eine Kaskade von Prozessen) zur für die Apoptose typischen Bläschenbildung und zum Zellabbau führt. (Capsasen sind Enzyme, die Proteine durch Hydolyse abbauen und bei der Apoptose eine besondere Rolle spielen.) Umgekehrt führte ein TNF von Korallen (AdTNF1) bei menschlichen Zellen zu einem signifikant häufigeren physiologisch hervorgerufenen Zelltod. TNF von Korallen und Menschen sind also austauschbar.
Beispiele wie der zelluläre Apoptose-Mechanismus zeigen einerseits ein erstaunliches Ausmaß an Konservierung eines zellulären Prozesses und seiner Bestandteile; in evolutionstheoretischer Interpretation muss ein Stillstand bezüglich der Evolution dieses Mechanismus von mindestens 550 Millionen Jahren angenommen werden. Andererseits werfen sie aber auch die Frage auf, wann und wie ein solcher Mechanismus überhaupt evolutiv entstand. Denn wenn er offenkundig nicht nennenswert veränderbar ist, wie sollen dann hypothetische Vorstufen in irgendeiner Weise funktional und damit existenzfähig sein? Die Daten zeigen einmal mehr: Abruptes Auftreten und dann weitgehend Konstanz eines zentralen biologischen Struktur- und Funktionsgefüges.
[Quistada SD, Stotlanda A et al. (2014) Evolution of TNF-induced apoptosis reveals 550 My of functional conservation. PNAS early ed, doi: 10.1073/pnas.1405912111]
|
|
PDF-Version
Eine im Juni 2014 veröffentlichte Pressemitteilung der Georg-August-Universität Göttingen lässt durch klare Aussagen aufhorchen: „Mond entstand durch Planetenkollision“ (Titel) und „Wissenschaftler bestätigen Impakthypothese“ (Untertitel). Die Pressemitteilung bezieht sich auf eine kürzlich in Science veröffentlichte Studie von Herwartz1 et al. (2014, Zitate unten in Übersetzung), in der erstmals eine minimale Differenz in der Sauerstoff-Isotopenzusammensetzung von Mond- und Erd-Gestein aufgezeigt wird. Dies wertet die Pressestelle der Universität als „sichere“ Bestätigung der Hypothese, nach welcher der Mond aus einer Kollision einer frühen Erde (Protoerde) und eines anderen Planeten bzw. Planetesimals (Theia genannt) entstand.
 |
| Abb. 1: Mond und Erde, ein historisches Bild aus dem Apollo-Programm (Foto: NASA). |
|
Für ihre aufwändige Dreifach-Sauerstoffisotopenanalyse untersuchten Herwartz et al. lunare Proben der Apollo-Missionen der Jahre 1969-72, terrestrische Proben (Minerale des Erdmantels) und Proben von Meteoriten (Enstatit2-Chondrite3). Als Maß dient das Verhältnis des seltenen Sauerstoffisotops 17O zum häufigsten Sauerstoffisotop 16O (Δ17O). Die Differenz zwischen den untersuchten terrestrischen und lunaren Gesteinen beträgt 12±3 ppm (Anteil pro Million). Dieses Ergebnis, so Herwartz et al., „(...) unterstützt die Kollisionshypothese der Entstehung des Mondes“. Die angenommene marsgroße Theia soll, so spekulieren die Autoren, eine Enstatit-chondritische Zusammensetzung gehabt haben (Δ17O-Differenz zu terrestrischen Proben 50 ppm).
Die Kollisionshypothese (giant impact hypotheses) ist die derzeit populärste Hypothese eines Hypothesen-„Wettstreites“ zur Entstehung des Mondes; ihre moderne Ausgestaltung geht auf Hartmann & Davis (1975) zurück. Eine Erde-Mond-„Verwandtschaft“ wird auf die im Wesentlichen identische chemische Zusammensetzung der beiden Himmelskörper zurückgeführt. Allerdings werden nach der Kollisionshypothese aufgrund der Isotopenheterogenität unter den Körpern des Sonnensystems Unterschiede erwartet; der theoretische Planet Theia soll deshalb eine von der Protoerde unterschiedliche Zusammensetzung gehabt haben, die sich bei der Kollision fraktionierend4 auf Protoerde und Mond ausgewirkt habe. Eine solche Fraktionierung wurde schon aufgrund von Computermodellen erwartet, konnte aber bislang nicht bestätigt werden. Die von Herwartz et al. ausgewiesene (minimale) Differenz wird nunmehr von der Presseabteilung der Universität Göttingen als entscheidendes Indiz für die Richtigkeit der Entstehungshypothese gewertet. Die Autoren drücken sich allerdings vorsichtiger aus – „der Mond wurde möglicherweise durch eine katastrophische Kollision der Protoerde mit einem Planetesimal namens Theia gebildet“, – und sie räumen ein, dass eine alternative Erklärung durch spätere Einwirkung von außen durchaus plausibel ist.
 |
| Abb. 2: Künstlerische Darstellung einer Planeten-Kollision (Bild: NASA/JPL-Caltech) |
|
Die Interpretation der Daten wirft einige grundsätzliche Fragen auf: Zunächst, ob die an einigen Proben ermittelten Δ17O-Werte jeweils repräsentativ für Gesamt-Erde und Gesamt-Mond sind. Des Weiteren, inwieweit die untersuchten Gesteine die Ursprungszustände widerspiegeln. Beispielsweise wurde erst kürzlich festgestellt, dass das Mondgestein unerwartet hohe Wassergehalte aufzeigt (Hui et al. 2013), und es erscheint deshalb unklar, ob es die ursprüngliche Mondkruste repräsentieren kann. Und nicht zuletzt muss der kausale Zusammenhang hinterfragt werden. Denn eine tatsächlich unterschiedliche Isotopenzusammensetzung muss nicht zwangsläufig von einer Planetenkollision herrühren. Aus diesen Gründen kann man höchstens den Schluss ziehen, dass dieser Befund der Kollisionstheorie nicht widerspricht, von einer „Bestätigung“ kann aber nicht die Rede sein.
Grenzüberschreitende Anmerkung: Mit der Kollisionshypothese wird nicht nur die Entstehung des Mondes, sondern zugleich auch die der Erde fixiert. Eingebettet in das Denkmodell einer Entstehung des Sonnensystems durch das Zusammenziehen einer Gaswolke soll die Protoerde-Theia-Kollision das letzte große Kollisionsereignis einer Vielzahl vorausgegangener Kollisionen gewesen sein und damit das Ende einer Millionen Jahre andauernden Erdentstehung in der frühen Geschichte dieses Sonnensystems markieren. In der Bibel (Genesis 1 und 2,1-2) finden wir eine Erklärung, die ohne Kollisionsmodell auskommt: Erde und Mond sind separate Schöpfungswerke in einer klar ausgewiesenen (Schöpfungs-) Reihenfolge. Gott sprach und es geschah. Dazu braucht es keine kosmische Evolution und keine langen Entstehungszeiten. Trotz seiner Kürze enthält der Schöpfungsbericht auch im Hinblick auf die Entstehung von Erde und Mond für den Menschen essentielle Detail-Informationen.
Anmerkungen: 1 Leitautor nunmehr Universität zu Köln; 2 Mineral der Pyroxen-Gruppe, Mg2[Si2O6]; 3 hauptsächlich aus Chondren (Silikatkügelchen, Si-O-Verbindungen) bestehend; 4 zu unter-schiedlichen Anteilen aufteilend.
[Georg-August Universität Göttingen, Pressemitteilung Nr. 131 vom 6. Juni 2014, Mond entstand durch Planetenkollision, www.uni-goettingen.de/de/3240.html?cid=4809; Hartmann WK & Davis DR (1975) Satellite-sized planetesimals and lunar origin. Icarus 24, 504-515; Herwartz D, Pack A, Friedrichs B & Bischoff A (2014) Identification of the giant impactor Theia in lunar rocks. Science 344, 1146-1150; Hui H, Peslier A, Zhang Y & Neal CR (2013) Water in lunar anorthosites and evidence for a wet early moon. Nature Geoscience 6, 177-180.]
Anmerkungen
- 1 Leitautor nunmehr Universität zu Köln; 2 Mineral der Pyro-xen-Gruppe, Mg2[Si2O6]; 3 hauptsächlich aus Chondren (Silikatkügelchen, Si-O-Verbindungen) bestehend; 4 zu unterschiedlichen Anteilen aufteilend.
|
|
|
In einem jurassischen Granit bei Rubiku (60 km nordöstlich Tirana, Albanien), im Mirdita-Ophiolith-Komplex, sind von Kryza & Beqiraj (2014) zwei unterschiedliche, aber in sich homogene Zirkon1-Populationen2 entdeckt worden; die interpretierten Isotopenalter betragen im Mittel 1.827 ± 17 und 169 ± 2 Millionen [radiometrische] Jahre. Die älteren Zirkone sollen von einem magmatischen Zirkon-Kristallisationsereignis herrühren. Sie waren „dann“, so die Autoren, zu einem überraschend großen Teil in der granitischen Magma enthalten, das bei der Platznahme des Ophioliths3 zur Kristallisation der jüngeren Zirkone führte (Bajocian-Stufe, jüngere Mitteljura).
Im Konzept der Plattentektonik werden Ophiolithe als Fragmente ozeanischer Lithosphäre interpretiert. Sie sind Kernbausteine in der Rekonstruktion der geodynamischen Prozesse sowie der Paläogeographie. Die Ophiolith-Abfolge im zentralen Albanien gehört zu dem mesozoischen Ophiolith-Komplex des alpidischen Orogensystems4 der Dinariden-Helleniden. Die Studie galt der Genese des Granitkomplexes als Teil der obduzierten, zerstückelten Ophiolith-Abfolge, insbesondere der zeitlichen Einordnung von Obduktion5 und Platten-Kollision. Mit dem (überraschenden) Vorkommen zweier altersunterschiedlicher Zirkon-Populationen hat die Aufgabenstellung allerdings deutlich an Komplexität gewonnen.
Kryza & Beqiraj diskutieren (vorsichtig) mögliche geodynamische Szenarien (mittelozeanischer Rücken, MOR; Supra-Subduktionzone, SSZ, Akkretions- und Kollisions-Situationen) und die sich daraus ergebenden paläotektonischen Implikationen. Heterogene Zirkon-Zusammensetzungen sind beispielsweise auch vom Mittelatlantischen Rücken bekannt (u. a. Pilot et al. 1998, Bortnikov et al. 2008). Sie kommen zu dem Schluss, dass die betrachteten Hypothesen möglicher geodynamischer Szenarien keine zufriedenstellenden Antworten, nicht einmal zu einigen wesentlichen Fragen, geben. Die Präsenz alter Zirkone in wesentlich jüngeren Gesteinen innerhalb ophiolithischer Abfolgen laufe aber geodynamischen Modellen zuwider, die eine Interaktion ozeanischer Lithosphäre mit kontinentaler Kruste vorsehen. Letztendlich können sie – nach eigenen Angaben – auf Basis der verfügbaren und limitierten Daten die Präsenz der alten Zirkone nicht erklären; es bedarf also weiterer Studien.
Bortnikov et al. (2008) berichteten zuvor von ungewöhnlich alten (etwa 100 bis 330 und sogar etwa 2.230 Millionen [radiometrische] Jahre) und jungen (etwa 1,2 bis 1,4 Millionen [radiometrische] Jahre) Zirkonen in denselben gabbroiden (basaltischen) Gesteinsproben aus der Markov-Tiefe des Mittelatlantischen Rückens. Das plattentektonische Konzept aber, so die Autoren, erwarte in der Spreizungszone nur die Bildung neuer ozeanischer Kruste, also nicht das Vorkommen unterschiedlich alter Zirkon-Populationen dieser Streuung. Unter der Annahme, dass die alten Zirkone artfremd sind, schlagen Bortnikov et al. (2008) zur Lösung des vermeintlichen Widerspruchs einen spezifischen geodynamischen Prozess im ozeanischen Mantel vor (S. 865-6): Demnach soll eine partielle Aufnahme von Material unterschiedlichen Alters vom Friedhof („graveyard“) subduzierter Krusten-Fragmente durch das Material eines aus dem Kern-Mantel-Grenzbereich aufströmenden Plumes (Manteldiapir) erfolgen. Während des Aufstiegs soll sich das Plume- und Krusten-Material noch in einem festen Aggregatszustand befinden. Das Material schmelze schließlich, wenn der Plume-Kopf die Soliduskurve überschreite und beginne sich (sodann) über die ozeanische Lithosphäre auszubreiten. Zirkon, als das höchste Temperatur-Mineral, würde zuletzt schmelzen und sich in dem basaltischen Magma auflösen.
Diese Deutung führt zu der grundsätzlichen Frage, warum sich die im Mantel befindlichen alten Krustenfragmente auf einer langen Zeitskala noch nicht thermisch an das Mantel-Material angeglichen haben und samt den Zirkon-Mineralen „resorbiert“ worden sind. Das Vorhandensein alten Krustenmaterials lässt vielmehr auf eine kurze Zeitachse schließen; das Material (der Friedhof) ist noch vorhanden, weil es nicht ausreichend lange „aufgeheizt“ wurde. So betrachtet ist die Zirkon-Vielfalt nicht nur eine Herausforderung für geodynamische Modelle, sondern auch für die lange Zeitachse.
[Bortnikov NS, Sharkov EV, Bogatikov OA, Zinger TF, Lepekhina EN, Antonov AV & Sergeev SA (2008) Finds of young and ancient zircons in gabbroids of the Markov Deep, Mid-Atlantic Ridge, 5°24‘-5°02.2’N (Results of the SHRIMP-II U-PB dating): implications for deep geodynamics of modern oceans. Doklady Earth Sciences 421, 859-866. – Kryza R & Beqiraj A (2014) Palaeoproterozoic (1.83 Ga) zircons in a Bajocian (169 Ma) granite within a Middle Jurassic ophiolite (Rubiku, central Albania): a challenge for geodynamic models. Int. J. Earth Sci. (Geol. Rundschau) 102, 607-625. – Pilot J, Werner C-D, Haubrich F, Baumann N (1998) Palaeozoic and proterozoic zircons from the Mid-Atlantic Ridge. Nature 393, 676-679.]
Anmerkungen
- 1 Mineral (ZrSiO4), teilweise mit geringen Mengen von Uran und Thorium; 2 Population: eine Gruppe von Zirkonen gleicher Art und gleichzeitiger Entstehung; 3 submarine basische bis ultrabasische (SiO2-arme) magmatische Gesteinsserien; 4 Orogenese: Gebirgsbildung; 5 auf das Festland „geschoben“.
|
|
| |
