 |
Plastizität: Quelle für evolutionär Neues?
von Reinhard Junker
Studium Integrale Journal
17. Jahrgang / Heft 2 - November 2010
Seite 99 - 102
|
 |
Zusammenfassung: Eines der Kennzeichen der Lebewesen ist ihre Fähigkeit, je nach Umwelteinflüssen verschiedene Merkmale oder Merkmalsausprägungen auszubilden. Diese Eigenschaft wird als Plastizität bezeichnet und es stellt sich die Frage, wie sie entstanden ist, insbesondere wenn man bedenkt, dass Plastizität eine zukunftsorientierte Fähigkeit ist. Lebewesen sind sozusagen in gewissen Grenzen auf manche Eventualitäten eingestellt. Zukunftsorientierung ist aus einer Designer-Perspektive leicht zu verstehen, für einen natürlichen Selektionsprozess dagegen eine Herausforderung. Ledon-Rettig et al. (2008) haben Experimente mit Kaulquappen durchgeführt, die je nach zur Verfügung stehender Nahrung unterschiedlich geformte Körpergestalten (sog. Morphen) ausbilden. Ein Vergleich verschiedener Gattungen gibt Hinweise auf einen möglichen Entstehungsweg von Plastizität. Diese Experimente werden vorgestellt und bewertet.
| |
|
Das Phänomen und eine Idee
Eine der vielen erstaunlichen Eigenschaften der Lebewesen ist ihre Plastizität (Formbarkeit). Gemeint ist damit die Fähigkeit, je nach Umweltreizen während ihrer Individualentwicklung oder auch im Erwachsenenzustand unterschiedliche Merkmale oder Merkmalsausprägungen ausbilden zu können, ohne dass eine Änderung des Erbguts zugrunde liegt. Ein bekanntes Beispiel ist die Verdickung der Hornhaut an den Händen von Waldarbeitern oder der Fingerkuppen bei Gitarrenspielern. Plastizität ist ein universelles Phänomen im Organismenreich; praktisch alle Organismen sind mehr oder weniger plastisch (Schlichting & Pigliucci 1998, 315). Man spricht in diesem Zusammenhang auch von Umweltsensitivität. Das heißt: Die Lebewesen können in Abhängigkeit von den Umweltbedingungen verschiedene, im Erbgut „schlummernde“ (latente) Entwicklungsprogramme abrufen. Man unterscheidet kontinuierliche Plastizität (als Reaktionsnorm bezeichnet) und diskontinuierliche Plastizität, also Ausprägungen sprunghaft verschiedener Formen (dann als Polyphenismus* bezeichnet). Ein Beispiel für einen Polyphenismus ist die Sommer- und Winterform des Landkärtchen-Falters, deren eine Form orange und die andere Form dunkelbraun gefärbt ist.
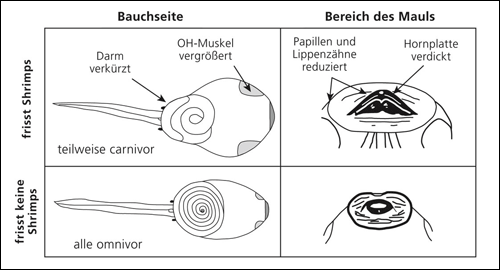 |
| Abb. 1: Vergleich des Baus von Kaulquappen der Gattung Spea, die Süßwassergarnelen (Shripms) fressen (oben) und solchen, die sich als Allesfresser ernähren (unten). (Nach Pfennig & Murphy 2000) |
|
Das Phänomen der Plastizität ist in der Frage nach der Entstehung evolutionärer Neuheiten in mancherlei Hinsicht interessant. Evolutionstheoretisch wird spekuliert, dass plastische Unterschiede Ausgangspunkte für evolutionäre Veränderungen sein könnten. Eine Idee besagt, dass in extremen Umwelten (bei Umweltstress) möglicherweise Extremvarianten ausgeprägt werden. Wenn sie anschließend durch Mutationen genetisch fixiert werden, können diese Änderungen im Erbgut beibehalten werden. Außerdem – so wird argumentiert – könnten sich gleichsam „im Schatten“ der Plastizität genetische Varianten ansammeln, die sich bei Umweltstress phänotypisch (im Erscheinungsbild) ausprägen könnten (Schlichting & Smith 2002, 203). Nicht-erbliche, plastische Modifikationen werden von einigen Forschern daher als wichtiger Schlüssel zum Verständnis von Makroevolution betrachtet.
Andererseits ist Plastizität ein auf Zukunft gerichtetes Kennzeichen. Die meisten Individuen einer Art benötigen ihre plastischen Möglichkeiten gar nicht, weil sie nicht mit entsprechenden stressenden Bedingungen konfrontiert werden. Plastizität beinhaltet Fähigkeiten, die bei Bedarf abgerufen werden können, also nur potentiell nützlich sind. Hier stellt sich die Frage, wie eine solche Fähigkeit evolutionär erworben werden könnte. Selektion kann nur den aktuellen Nutzen „bewerten“, nicht einen potentiellen.
Selektion kann ja immer nur einen bestimmten Zustand im Rahmen einer Modifikationsspanne bewerten. Wie soll sie bewerten, dass es gut wäre, mehrere Körpergestalten (Morphen*) für unterschiedliche Umwelten parat zu haben, falls sich diese im individuellen Leben verändern? Selektion kann sich ja auch nicht an verschiedene Umwelten erinnern, in denen ein Organismus in einer bestimmten Zeitspanne gelebt hat, um die passende Modifikationsspanne zu selektieren (stabile Umwelt: kleine Spanne, labile: große Spanne).
Aus einer Designer-Perspektive sind solche Fähigkeiten insofern leicht zu verstehen, als ein Designer die Zukunft gedanklich vorwegnehmen und seine Geschöpfe darauf einstellen kann.
Forschungen zur Entstehung von Plastizität stehen noch am Anfang. Nachfolgend soll ein Beispiel einer Studie zur Entstehung von Plastizität vorgestellt werden.
|
|
Das Experiment
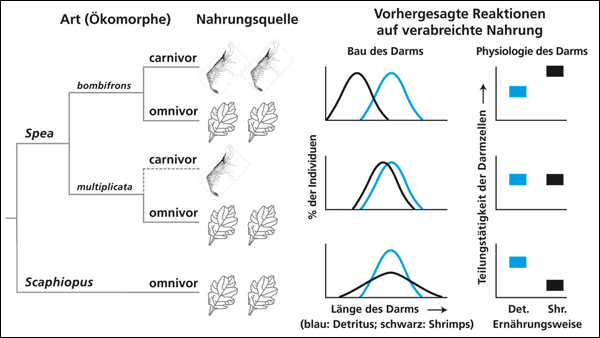 |
| Abb. 2: Links: Cladogramm und Ernährungsweise der im Text beschriebenen Arten. Rechts: Reaktionen der Darmlänge und der Darmphysiologie bei unterschiedlicher Ernährung. Näheres im Text. (Nach Ledon-Rettig et al. 2008) |
|
Ein interessantes Experiment zur Entstehung von Plastizität haben Ledon-Rettig et al. (2008) bei Kaulquappen durchgeführt. In der Gattung Spea gibt es eine Art, S. bombifrons, die einen ausgeprägten Polyphenismus zeigt: Wenn sie sich von Süßwassergarnelen (Shrimps) ernährt, bildet sie einen dickeren und verkürzten Darm aus, der Kopf ist größer und die Physiologie des Darms wird durch Erhöhung der Zellteilungsrate verändert (carnivore Form, Abb. 1). Ernährt sich diese Art dagegen mit Detritus, ist der Darm lang und dünn, der Kopf kleiner und die Zellteilung im Darm wird nicht verstärkt (omnivore Form). Eine zweite Art, S. multiplicata, kann sich zwar auch von Garnelen und von Detritus ernähren, ist aber nicht so plastisch wie S. bombifrons. Die Kaulquappen dieser Art verändern bei carnivorer Nahrung kaum ihre Darmform. Bei Konkurrenz der beiden Arten kann S. bombifrons, wenn ein entsprechendes Nahrungsangebot vorliegt, effektiv auf Garnelen ausweichen und der Konkurrenz entgehen. Es handelt sich also um eine Konkurrenzvermeidung durch Kontrastverstärkung. Die carnivore Form ist somit in kurzlebigen Wasserstellen begünstigt, da es dort viele Garnelen gibt und ihr schnelles Wachstum zum Überleben hilft. Omnivore Kaulquappen dagegen überleben besser in langlebigeren Wasserstellen, weil sie durch das langsamere Wachstum die Metamorphose in einer besseren Konstitution erreichen.
Ledon-Rettig et al. führten Versuche aus, um herauszufinden, wie die Plastizität bei S. bombifrons entstanden sein könnte. Sie untersuchten die nahe verwandte Gattung Scaphiopus, die nach phylogenetischen Studien als Vorläufer von Spea angesehen wird. Diese Gattung ist omnivor, kann aber auch carnivor überleben, falls nur Garnelen als Nahrungsquelle zur Verfügung stehen. Die Forscher stellten fest, dass sich bei Anwesenheit von Garnelen die Variabilität der Darm-Ausgestaltung in alle Richtungen erhöht, also sowohl auf eine Weise, die zur carnivoren Morphe von S. bombifrons (kurzer, dicker Darm) als auch in die andere Richtung führt (Abb. 2). Der Darm zeigt also eine Plastizität, aber keinen ausgeprägten Polyphenismus wie bei S. bombifrons; die Plastizität ist latent bereits vorhanden und wird durch eine ungewohnte Umwelt (neues Nahrungsangebot) zur Ausprägung gebracht. Dagegen wird die Teilungstätigkeit der Darmzellen durch die Anwesenheit nicht verstärkt; dieses Merkmal ist bei Scaphiopus nicht plastisch.
Ledon-Rettig et al. interpretieren diesen Befund so, dass der Polyphenismus bei S. bombifrons sich aus einer zunächst in verschiedene Richtungen sich ausprägenden Plastizität entwickelt haben dürfte. Ausgangspunkt ist also eine schon vorhandene, aber nur allgemeine Plastizität. Daraus könnte sich durch Fixierung bestimmter Ausprägungen innerhalb der Plastizitätsgrenzen der Polyphenismus von S. bombifrons entwickelt haben.
Man kann also folgendes Szenario vermuten: Zunächst ist eine latente (verborgene) Fähigkeit zur Carnivorie vorhanden; das Merkmal „Art der Nahrungsaufnahme“ ist also plastisch. Auf dieser Basis erfolgte eine genetische Akkommodation*, d. h. die Umweltsensitivität wurde verändert, wodurch die angebotene Nahrung (Garnelen) besser genutzt werden kann (Ledon-Rettig et al. 2008). Die Art S. multiplicata, deren Plastizität verringert ist, könnte daraus durch genetische Assimilation entstanden sein. Das heißt, eine bestimmte Ausprägung innerhalb der Plastizitätsspanne wurde (weitgehend) fixiert, wodurch die Plastizität (bzw. die Umweltsensitivität) verringert wurde. Also kurz zusammengefasst (vgl. Abb. 2): Unspezifische Plastizität (wie bei Scaphiopus) Æ Polyphenismus (wie bei Spea bombifrons) Æ Assimilation der einen polyphenen Morphe (wie bei S. multiplicata).
|
Genetische Akkommodation: Vorgang, durch den die Umweltsensitivität eines Merkmals verändert wird und diese Änderung auch durch Änderung des Erbguts fixiert wird. Plastizität: Änderungen in Organismen infolge von Umweltreizen (Schlichting & Smith 2002, 190). Wenn die umweltbedingten Modifikationen graduell sind, spricht man von „Reaktionsnormen“; sind die unterschiedlichen Ausprägungen dagegen sprunghaft verschieden, handelt es sich um einen Polyphenismus („Vielgestaltigkeit“). Polyphenismus: diskontinuierliche Plastizität, Ausprägung sprunghaft verschiedener Formen innerhalb einer Art. Morphe: Ausprägung einer bestimmten Gestalt innerhalb einer plastischen Art. |
|
|
Bewertung
Ledon-Rettig et al. (2008) stellen fest, dass kein bekannter Mechanismus neue Umwelten vorhersehen und entsprechende Anpassungen bereitstellen kann. Dies wird in ihrem Experiment insofern bestätigt, als die plastische Veränderung des Darms von Scaphiopus in keine bestimmte Richtung ging, als der neue Umweltreiz „ das Angebot von Garnelen als Nahrungsquelle „ auftrat. Die physiologischen Eigenschaften des Darms wurden in dieser vermuteten Vorläuferart zudem kaum verändert. In einem insgesamt engen Rahmen ist mit diesem Experiment die Entstehung eines Polyphenismus, ausgehend von einer unspezifischen Plastizität, wahrscheinlich gemacht. Die Änderung der Physiologie des Darms, nämlich die erhöhte Teilungstätigkeit der Darmzellen, müsste somit nachfolgend optimiert worden sein. Woher die bei Scaphiopus vorhandene Plastizität des Darms kommt, wird mit diesem Experiment nicht geklärt. Eine ungerichtete Plastizität ist als Ausgangssituation vermutlich notwendig.
 |
Eines der bedeutendsten ungelösten
Probleme der Biologie ist das
Verständnis, wie neue, komplexe
Phänotypen entstehen, sowohl in der
individuellen Entwicklung als auch
in der Stammesgeschichte. |
 |
|
Die Schlussfolgerung der Autoren steht und fällt mit der phylogenetischen Position von Scaphiopus, die gegenüber der polyphenen Gattung Spea als ursprünglicher bestimmt wurde. Sollte sich durch weitere Befunde herausstellen, dass Scaphiopus von einer polyphenen Gattung abzuleiten ist, müsste die ungerichtete Plastizität bei Scaphiopus als Verlust eines Polyphenismus gewertet werden. Weitere Studien an verwandten Gattungen können die verschiedenen Deutungsmöglichkeiten stärken oder schwächen.
Das Beispiel der Plastizität des Baus und der Physiologie des Darms der Kaulquappen ist nicht geeignet, die Entstehung neuer Strukturen plausibel zu machen. Die Idee, dass versteckte Plastizitätsspannen Potential für evolutionäre Neuheiten bereithalten können, ist durch die Änderungen in der mutmaßlichen Mikroevolution von Scaphiopus zu Spea nicht gedeckt. Ledon-Rettig et al. (2008) beginnen ihren Artikel mit dem Satz: „Eines der bedeutendsten ungelösten Probleme der Biologie ist das Verständnis, wie neue, komplexe Phänotypen entstehen, sowohl in der individuellen Entwicklung als auch in der Stammesgeschichte.“
Dieser Satz behält auch trotz ihrer interessanten Experimenten seine Gültigkeit.
|
|
Literatur
- Ledon-Rettig CC, Pfennig DW & Nascone-Yoder H (2008)
- Ancestral variation and the potential for genetic accommodation in larval amphibians: implications for the evolution of novel feeding strategies. Evol. Dev. 10, 316-325.
- Pfennig DW & Murphy PJ (2000)
- Character displacement in polyphenic tadpoles. Evolution 54, 1738-1749.
- Schlichting CD & Pigliucci M (1998)
- Phenotypic Evolution: A Reaction Norm Perspective. Sunderland, Mass.
- Schlichting CD & Smith H (2002)
- Phenotypic plasticity: linking molecular mechanisms with evolutionary outcomes. Evol. Ecol. 16, 189-211.
|
|
|  |
