 |
|
von Reinhard Junker
Studium Integrale Journal
17. Jahrgang / Heft 2 - November 2010
Seite 59 - 67
|
 |
Zusammenfassung: Im ersten Teil dieses Artikels (Stud. Int. Journal 17, 21-28) wurden Hypothesen zur Entstehung des Schildkrötenpanzers vorgestellt und diskutiert. Außerdem wurden die isolierte systematische Stellung und das plötzliche Erscheinen der Schildkröten in der Fossilüberlieferung thematisiert. Im nun folgenden zweiten Teil werden neue Fossilfunde vorgestellt und im Rahmen der konkurrierenden Panzer-Entstehungs-hypothesen diskutiert. Die in den Jahren 2008 und 2009 veröffentlichten fossilen Gattungen Odontochelys und Chinlechelys besaßen Merkmale, aus denen Hinweise auf ihre hypothetische Evolution entnommen werden können. So besaß Odontochelys keinen verknöcherten Rückenpanzer, und der Panzer von Chinlechelys war sehr dünn und nur teilweise mit den Rippen verwachsen. Es ist aber in beiden Fällen nicht sicher, ob es sich dabei um phylogenetische Primitivmerkmale handelt. Odontochelys könnte auch eine spezialisierte wasserlebende Gattung gewesen sein. Und die fossilen Reste von Chinlechelys sind möglicherweise zu spärlich, um sichere Schlussfolgerungen zu erlauben.
Anschließend wird eine Reihe von Fragen zur Evolution der Schildkröten zusammenfassend diskutiert: Ontogenetische* Befunde und die Bedeutung der Carapaxfalte*, die uneinheitliche systematische Stellung der ältesten Fossilfunde, molekulare Daten und der Fossilbefund, einzigartige Merkmalskombinationen und Konvergenzen*, Probleme des Umbaus zum Panzer und die Ontogenese als fragwürdiges Modell für die Phylogenese*.
Abschließend werden einige spekulative Ideen für die Möglichkeit einer nicht-evolutionären Deutung der Fossilüberlieferung der ältesten Schildkröten erläutert.
| |
|
Neue Fossilfunde
Fassen wir den Kenntnisstand über die ältesten Schildkrötenfossilien bis 2007 kurz zusammen. Der Schildkröten-Bauplan erscheint fossil ohne dokumentierte Bindeglieder; vermittelnde Formen fehlen unabhängig von der systematischen Stellung der Schildkröten. Die frühen Fossilien besitzen aber eine Reihe von „Primitivmerkmalen“, die sie mit anderen Reptiliengruppen teilen. So schützten sie ihren Kopf nicht durch Rückziehen in den Panzer (Halswender oder Halsberger), sondern mit Nackenstacheln oder mit einem Fortsatz des Carapax. Vom Fossilbericht her und aufgrund phylogenetischer Analysen (Joyce 2007) spricht einiges für eine terrestrische* Entstehung der Schildkröten. Hinsichtlich der kausalen Erklärung der Entstehung des Schildkrötenpanzers stehen sich die transformistische und die emergentistische Hypothese gegenüber. Zur Erinnerung: Nach der transformistischen Sicht entstand der Panzer aus Hautknochen unter der Wirkung der natürlichen Selektion. Nach dieser Hypothese kam es bei den Schildkröten zu einer Fusion von Hautknochen mit den Rippen und Wirbelfortsätzen. Nach der emergentistischen Hypothese ist der Panzer dagegen durch Veränderungen in der ontogenetischen Entwicklung entstanden. Durch Neuverschaltungen von Steuergenen soll sich unabhängig von selektionsgesteuerten Veränderungen die neue Struktur des Panzers gebildet haben. Nach den bis dato bekannten Befunden schien die emergentistische Hypothese trotz großer Erklärungslücken plausibler.
Bezüglich des Zeitpunkts der phylogenetischen Abzweigung der Schildkrötenlinie bestand zwischen dem Fossilbefund und den molekularen Befunden eine deutliche zeitliche Diskrepanz (ca. 50 Millionen Jahre).
Im Jahr 2008 wurden nun zwei neue bedeutende Fossilfunde gemacht. Sie sollen zunächst beschrieben und anschließend bezüglich ihrer Bedeutung für die oben aufgeworfenen Fragen (Ent-stehung auf dem Land oder im Wasser; transformistisch oder emergentistisch) bewertet werden.
Odontochelys
Li et al. (2008) berichten von Fossilfunden einer Schildkröte, die einen Zwischenschritt in der Evolution des Panzers dokumentieren soll. Die neue Art Odontochelys semitestacea (Abb. 1) wurde in Schichten in der südwestchinesischen Provinz Guizhou in Küsten-Ablagerungen entdeckt, die mit datierten 220 Millionen Jahren noch etwas älter sind als die bislang älteste Schildkröte Proterochersis (ca. 214 Millionen Jahre; s. o.). Der Artname bedeutet sinngemäß „Halbpanzerschildkröte mit Zähnen“, womit zwei wesentliche Unterschiede zu heutigen Schildkröten angedeutet sind. Zum einen waren Ober- und Unterkiefer des Tieres bezahnt und hatten keine schnabelartigen Kieferleisten wie heutige Schildkröten. Zum anderen war zwar der Bauchpanzer (Plastron) des etwa 40 cm großen Tieres voll entwickelt, der Rückenpanzer (Carapax) bestand aber nur aus Neuralplatten; die rückenseitigen Rippen waren lediglich verbreitert. Hautknochen waren nicht ausgebildet. Li et al. (2008) schließen daraus, dass sich der bauchseitige Panzer vor dem Rückenpanzer entwickelt hat und dass der erste Schritt der Entstehung des Rückenpanzers in der Verknöcherung von Neuralplatten und Verbreiterung der Rippen bestand. Diese Abfolge stimmt mit dem Verlauf der frühen Embryonalentwicklung der heutigen Schildkröten überein. Gegen die Deutung, es handle sich um ein jugendliches Exemplar, spricht nach Li et al. (2008, 499) die Verschmelzung von Sprungbein (Astragalus) und Fersenbein (Calcaneum). Nagashima et al. (2009) vermuten, dass in Odontochelys die Carapaxfalte* („carapacial ridge“, CR) nur an der Seite des Körperstamms ausgebildet und am vorderen und hinteren Ende unvollständig war. Erst deren Vervollständigung bewirkte das veränderte Rippenwachstum. Li et al. (2008) stellen demnach die Hypothese auf, dass sich der Schildkrötenpanzer in zwei Etappen gebildet hat, zuerst das Plastron, danach der Carapax, während man bislang davon ausgegangen war, dass sich beide Panzerteile zusammen entwickelt hätten (Maisch 2009, 201). Entsprechend soll sich die CR in zwei Etappen gebildet haben (Nagashima et al. 2009, 196). Im Wasser wäre eine solche Abfolge insofern plausibel, als ein Bauchpanzer vor Feinden von unten (vor lauernden Räubern) schützen konnte, während ein Schutz von oben weniger erforderlich gewesen wäre.
 |
| Abb. 1: Odontochelys semitestacea, specimen number IVPP V13240. Foto: Mr. Wei Gao. Abdruck mit freundlicher Genehmigung von Chun Li. |
|
Bei Odontochelys liegt das Schulterblatt vor dem ersten Rippenpaar, was einem ontogenetischen Stadium der heutigen Schildkröte Pelodiscus entspricht (Nagashima et al. 2009). Dies kann als Indiz dafür gewertet werden, dass Odontochelys als Übergangsform passen würde.
Der Interpretation von Li et al. stellen Reisz & Head (2008) in einem Kommentar jedoch eine andere entgegen: Lange, verbreiterte Rippen sind Bestandteile des Rückenpanzers aller Schildkröten. Deren Vorkommen bei Odontochelys sei ein Hinweis darauf, dass das embryonale Gewebe, welches die Bildung des Carapax kontrolliere, ebenfalls vorhanden gewesen sei. Ebenso sei die Verbindungsbrücke zwischen Carapax und Plastron ausgebildet, was ebenfalls für die Anwesenheit eines Carapax spreche. Zusammengenommen spreche das für die alternative Interpretation, dass ein Carapax doch ausgebildet gewesen sei, jedoch einige seiner Teile nicht verknöchert gewesen seien. Diese Interpretation von Odontochelys führe zur Möglichkeit, dass dessen Panzer nicht primitiv, sondern stattdessen eine spezialisierte Anpassung durch einen sekundären Verlust gewesen sei (Reisz & Head 2008, 451), also keine Ausprägung, die einem frühen Ontogenesestadium entspricht. Eine Reduktion der Hautknochen ist bei wasserlebenden Schildkröten verbreitet; die Deutung einer Spezialisierung passt also zum vermutlichen Lebensraum von Odontochelys im küstennahen Wasser. Diese Ausprägung könne durch ein Stehenbleiben der Entwicklung auf einem jugendlichen Stadium (sog. Pädomorphose) zustande gekommen sein. Eine schwache Panzerung kommt bei vielen fossilen und rezenten Formen des Meeres und des Süßwassers vor (Maisch 2009). Wegen eindeutiger Merkmale des Erwachsenenzustands, der Körpergröße und Fusion von Fußwurzelelementen „ist auszuschließen, dass die zwei Fossilien Jungtiere waren“ (Maisch 2009, 201).
Allerdings passt die Bezahnung, die evolutionstheoretisch als ursprüngliches Merkmal gelten muss, nicht ohne weiteres zu dieser Deutung. Denn wenn Odontochelys als eine spezialisierte Schildkrötenart interpretiert wird, wäre eher zu erwarten, dass sie wie die anderen Schildkröten Hornplatten statt Zähne besitzen. Außerdem gelten das Basipterygoidgelenk zwischen Gaumen und Hirnschädelbasis, das den Gaumen wie bei Proganochelys beweglich machte, und die Flügelbeine (Pterygoide) im Gaumenbereich mit vermutlich ausgebildeten Querfortsätzen, die bei Proganochelys reduziert sind, als Primitivmerkmale. Allerdings können Merkmale auch mosaikartig kombiniert sein; eine spezialisierte wasserlebende Art kann bezahnt gewesen sein. Einerseits ist der Rückenpanzer von Odontochelys mit keinen frühontogenetischen Stadien heutiger Schildkröten vergleichbar und sein Bau weicht von dem anderer aquatischer* Schildkröten ab. Andererseits ist das Plastron von Odontochelys gut verknöchert und „lässt sich nicht von dem anderer Schildkröten unterscheiden“ (Maisch 2009, 201). Die Rippen sind wie bei heutigen Schildkröten nicht ventral stark gebogen, sondern seitwärts abstehend und verkürzt. Wird das Fehlen des Carapax bei Odontochelys als ursprünglich interpretiert, stellt sich die Frage nach den Selektionsdrücken, die die Entstehung einer Vorstufe des Carapax wie z. B. seitwärts stehender Rippen begünstigen konnten. Im Falle einer sekundären Rück-bildung erübrigt sich diese Frage, da sich ein Verlust viel leichter ereignen kann als ein Neuerwerb.
Wie bei der ebenfalls obertriassischen voll gepanzerten Gattung Proganochelys ist das Schädeldach anapsid (kein Schläfenfenster), obwohl – wie bereits ausgeführt – nach phylogenetischen Studien die Schildkröten zu den Diapsiden zu stellen sind, womit das Fehlen der Schädelfenster als sekundär zu deuten ist. Dass bereits die ältesten Schildkrötenfunde einen anapsiden Schädelbau haben, passt nicht zur phylogenetischen Stellung als späte Amnioten-Linien. „Die Fossilien geben damit nach wie vor den Befürwortern einer Abstammung der Schildkröten von primär anapsiden Vorfahren Schützenhilfe“ (Maisch 2009, 201). Insgesamt besitzt Odontochelys also ein Merkmalsmosaik, das nicht leicht in ein stammesgeschichtliches Schema eingeordnet werden kann. Wie in vielen anderen Fällen zeigt sich, dass Mosaikformen nicht ohne weiteres als evolutive Übergangsformen interpretierbar sind.
Die alternative Interpretation der Befunde durch Reisz & Head würde die Frage nach dem Ursprung der Schildkröten unverändert offen lassen und sogar verschärfen: Denn demnach wäre ausgerechnet eine spezialisierte Form die bislang älteste. Die Funde von Odontochelys machen auch beispielhaft deutlich, wie schwierig die Interpretation fossiler Funde ist.
Zur Frage, ob Schildkröten auf dem Land oder im Wasser entstanden sind, kann Odontochelys an-gesichts der verschiedenen Interpretationsmöglich-keiten keine sichere Auskunft geben. Die Gattung war zwar mit großer Wahrscheinlichkeit wasserlebend, aber eine sekundär aquatische Lebensweise kann nicht ausgeschlossen werden.
Die Merkmalskombination von Odontochelys passt im Rahmen des Evolutionsparadigmas eher zur Emergenzhypothese, da der Fund – evolutions-theoretisch interpretiert – nicht zur Vorstellung eines allmählich rundum entstehenden Panzers passt.
Chinlechelys
Nachdem mit der Entdeckung von Odontochelys die Waage überraschend zugunsten einer Entstehung der Schildkröten im Wasser ausgeschlagen ist, scheint ein weiterer kurz darauf veröffentlichter ähnlich alter Fund diese Deutung wieder „massiv in Frage“ zu stellen (Maisch 2009, 201). Joyce et al. (2009) beschreiben Bruchstücke einer auf 215 Millionen Jahre datierten fossilen Schildkröte Chinlechelys tenertesta aus der Obertrias von New Mexico, bei der Panzer und Rippen nicht miteinander verschmolzen waren. Erhalten sind allerdings nur wenige Osteodermen (Hautverknöcherungen) des Hals- und Schwanzbereichs und Elemente des Panzers (Teile des Carapax und Hypoplastrons), weshalb weitergehende Schlussfolgerungen nur vorbehaltlich weiterer Erkenntnisse durch vollständigere Funde gemacht werden können.
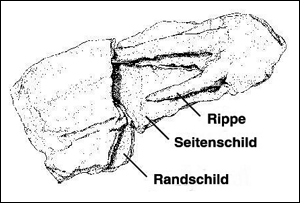 |
| Abb. 2: Ventrale (bauchseitige) Ansicht eines Bruchstücks des Carapax con Chinlechelys, bestehend aus zwei Seitenschilden, zwei Rippen und einigen Randschilden. Die Rippen haben nur eine schwache Verbindung mit den Costalia. (Nach Joyce et al. 2009; fig. 1g, Abdruck mit freundlicher Genehmigung von der Royal Society London) |
|
Chinlechelys tenertesta besitzt Nackenstachel, die aus mehreren Osteodermen aufgebaut sind. Nach Auffassung von Joyce et al. deuten sie an, dass die ersten Schildkröten von Reihen aus schützenden Strukturen (Schilden etc.) der Haut bedeckt waren. Wesentlich bedeutender sei jedoch, dass die primitiven, vertikal orientierten dorsalen Rippen der neuen Schildkröte nur eine schwache Verbindung mit den darüber liegenden Seitenschilden des Carapax aufweisen (Abb. 2). Das weise darauf hin, dass es sich hier um zwei unabhängig voneinander verknöchernde Strukturen handelt. Die Carapax-Knochenelemente sind sehr dünn (bis maximal drei Millimeter). Diese Beobachtungen an Chinlechelys unterstützen die Hypothese, dass der ursprüngliche Schildkrötenpanzer nur aus Schutzstrukturen der Haut bestand, und zwar rundum, der dann erst später mit den Rippen und Wirbeln des Innenskeletts verschmolzen ist und es sich somit beim Schildkrötenpanzer nicht um eine „Neuerfindung“ handelt, sondern dass er sich eher aus einer kontinuierlichen Vorläuferlinie entwickelt hat. Der Fund passt also – anders als Odontochelys – eher zur klassischen transformistischen Entstehungs-hypothese. Die lose Verbindung zwischen den Rippen und den Hautknochen, die sehr dünne Panzerung und der „beginnende doppelte Kontakt der dorsalen Rippen mit den Centra der Wirbel“ können gut als intermediäre Ausprägungen zwischen nichtgepanzerten Vorläufern und vollgepanzerten Schildkröten interpretiert werden (Joyce et al. 2009, 510f.). Die zahlreichen Hautknochen im Nacken- und Schwanzbereich seien ein Argument dafür, dass der Panzer ein „Komposit aus Hautknochen, Wirbeln und Rippen“ darstellt (Maisch 2009, 201).
Die für die Beurteilung wesentlichen Panzerelemente (Seiten- und Wirbelsäulenschilde, Costalia und Neuralia) sind also keine Auswüchse (Verbreiterungen) der Knochen des Innenskeletts (Rippen, Wirbel), wie einige embryologische Untersuchungen nahelegen. Joyce et al. betrachten damit die embryologischen Befunde, wonach der Panzer direkt aus den Rippen entsteht, als nicht maßgeblich. Darauf kommen wir weiter unten zurück.
Unklar ist, welche funktionale Bedeutung die veränderte Lage der Rippen hat, wenn diese nur lose mit den Hautknochen verbunden sind. Die Möglichkeit, dass es sich um ein frühes ontogenetisches Stadium handelt, halten Joyce et al. (2009, 509) für eher unwahrscheinlich. Sie merken an, dass das relative ontogenetische* Alter zwar schwer zu bestimmen sei, sie sind sich aber ziemlich sicher, dass die besondere Orientierung und Platzierung der Rippen und die Platzierung sowie geringe Dicke des Panzers keine juvenilen* Ausprägungen sind. Zusätzliches Fundmaterial könne einen Test auf diese Interpretation liefern.
Dieser Fund spricht nun wiederum für eine terrestrische* Entstehung der Schildkröten und für das klassische Modell und lässt sich mit Odontochelys nicht in eine Reihe bringen, es sei denn man interpretiert das Fehlen der Rückenpanzerung bei Odontochelys doch als Rückbildung. Andernfalls müssten Chinlechelys und Odontochelys als Vertreter deutlich verschiedener evolutiver Linien interpretiert werden, was auf eine mindestens biphyletische* Entstehung der Schildkröten hinausliefe.
|
abgeleitetes Merkmal: Merkmal, das als „höherentwickelt“ eingestuft wird. AER, Apikalfalte („apical ectoder-mal ridge“): Das körperferne Ende (Ektodermkappe) der Extremitätenknospe. Das Ektoderm an der Spitze der Knospe verdickt sich und bildet eine spezialisierte Struktur, die Apikal-leiste oder Apikalfalte. Amnioten: Landwirbeltiere, deren Embryonen sich in einer mit Fruchtwasser gefüllten Amnionhöhle entwickeln. aquatisch: im Wasser lebend. biphyletisch: auf zwei unabhängigen Abstammungslinien, zweimal unabhängig entstanden. Carapaxfalte: Aufwölbung, die sich oberhalb der Extremitätenknospe in Längsrichtung auf der Körperflanke bildet (vgl. Abb. 3). Duplikation: Verdopplung. Epithel: Deckgewebe. juvenil: im Jugendalter. Konvergenz: gleichartige Entwicklung ähnlicher Strukturen und Funktionen einzelner Organe nicht abstam-mungsmäßig verwandter Organismen. Mesenchym: embryonales Bindegewebe. Morphologie: Lehre vom Bau und der Gestalt der Organismen. Ontogenese: Individualentwicklung von der befruchteten Eizelle an. Osteodermen: Hautverknöcherungen. Phylogenese: Stammesgeschichte. Schwestergruppen: Zwei Taxa (Gruppen), die auf eine nur ihnen ge-meinsame Stammart zurückgeführt werden können. terrestrisch: auf dem Land lebend.
|
|
|
Zusammenfassende Diskussion
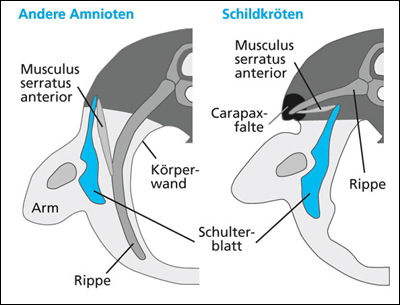 |
| Abb. 3: Lage der Carapaxfalte und ungewöhnliche Lage des Schulterblatts und der Rippen bei Schildkröten im Vergleich zu anderen Amnioten. (Nach scienceblogs.com) |
|
Ontogenetische Befunde und die Bedeutung der Carapaxfalte
Wie entstand die Carapaxfalte („carapacial ridge“, CR) und wie entstanden die Interaktionen mit der CR, die zur Bildung des Panzers führten? Im ersten Teil dieses Artikels wurde die Bedeutung der CR für die ontogenetische Bildung des Panzers erläutert. Die CR, eine Aufwölbung aus Ektoderm und Mesoderm, die sich oberhalb der Extremitätenknospe in Längsrichtung auf der Körperflanke bildet, ist eine einzigartige Struktur bei den Schildkröten, und soll zu einer neuen Epithel*-Mesenchym*-Interaktion geführt haben, die das Rippenwachstum beeinflusst. Durch die Wechselwirkung mit der CR biegen die Rippen zur Seite ab und der Schultergürtel positioniert sich innerhalb des Brustkorbs (Abb. 3). Die CR wird als eine Art Schlüssel-Organanlage betrachtet, die eine notwendige Voraussetzung für die Bildung des ungewöhnlichen Schildkrötenbauplans darstellt. Ebenfalls im ersten Artikelteil (Abschnitt „Evolutionäre Folgerungen“) wurde die Hypothese der Entstehung des Schildkröten-Panzers als Folge einer Mega-Duplikation (Verdopplung) erläutert. Die Duplikation der Randleiste („apical ectodermal ridge“, AER) der Extremitätenknospe soll zu einer Aufwölbung aus Ektoderm und Mesoderm und damit zur CR geführt haben, was wiederum ein verändertes Rippenwachstum zur Folge hatte und mit der Bildung des Panzers korreliert sei. Die CR wird nach dieser Hypothese als Schlüsselneuheit interpretiert, die zur Entstehung des Schildkrötenbauplans führte.
Damit wird aber nur eine notwendige, jedoch bei weitem nicht hinreichende Voraussetzung für die Entstehung des Schildkrötenpanzers geliefert. Denn der Weg zur „Installation“ der CR auf dem Wege einer Duplikation ist vollkommen unklar. Eine „Mega-Duplikation“ (mit den im ersten Teil diskutierten Einschränkungen) kann nur dann funktional sein, wenn viele Wechselwirkungen aufeinander abgestimmt werden; es sind bei einem solchen Vorgang sonst schwere Missbildungen zu erwarten. Wie eine solche vielfache Abstimmung von Einzelteilen vor sich gehen soll, also wie z. B. die Duplikation der AER das veränderte Rippenwachstum und die anderen Veränderungen hervorgebracht haben soll, die wiederum zur Bildung des Panzers geführt haben, darüber wird keine Rechenschaft abgegeben. Daher steht die Idee, mit der evolutiven Neubildung der CR eine schnelle und sehr umfangreiche Neubildung plausibel machen zu können, auf sehr schwachen Füßen. Die hypothetischen evolutiven Vorgänge müssten konkret beschrieben werden und es müsste eine Analyse erfolgen, welche Wechselwirkungen berücksichtigt werden und welche neuen Abstimmungen zwischen den Geweben und den ontogenetischen Entwicklungswegen erfolgen müssten. Gilbert et al. (2001, 56) argumentieren zwar, dass die Wechselwirkung zwischen Epithel* und Mesenchym* am Beginn der Carapax-Entwicklung die Basis für Hypothesen einer schnellen Evolution des Körperbauplans liefere, doch diese „Basis“ liefert allenfalls eine notwendige Voraussetzung für eine Erklärung, der hypothetische Prozess der Veränderung wird damit nicht aufgeklärt.
Dasselbe gilt für Überlegungen von Nagashima et al. (2005, 149). Diese Autoren betrachten die Verschiebung des Rippenwachstums in seitliche Richtung als Basis für veränderte Wechselwirkungen zwischen den Geweben, die zu den Hautknochen und verbreiterten Platten des Carapax führen. Eine kleine räumliche Veränderung in der Entwicklung (Heterotopie) könne zu einer größeren morphologischen Änderung führen. Auch hierbei handelt es sich nur um Begleiterscheinungen oder notwendige Voraussetzungen, denn eine Heterotopie für sich alleine kann keine Erklärung sein, da die verschiedenen Bauelemente koordiniert geändert werden müssen, was zahlreiche aufeinander abgestimmte Schritte benötigen würde. Auch Cebra-Thomas et al. (2005) beschreiben in ihren Modell nur notwendige Bedingungen bzw. Begleiterscheinungen der Panzerbildung.
Die alternative Erklärung einer allmählichen Entstehung des Panzers durch sukzessives Größerwerden von Hautknochen und ihr Verschmelzen mit Rippen und Wirbelteilen wirft auf der anderen Seite die Frage auf, wie es unter diesen Umständen zu den markanten Gemeinsamkeiten zwischen der CR und der AER kommen konnte (die im Abschnitt „Evolutionstheoretische Folgerungen“ des ersten Artikelteils dargestellt wurden). Rieppel (2009) nennt zwei Probleme der Transformationshypothese: Wie konnte es zur Fusion mit den Rippen kommen, da sie normalerweise in den Bauchraum wachsen? Und wie gelangt das Schulterblatt ins Innere des Brustkorbs? Nach der transformistischen Hypothese bewegte sich der Schultergürtel während der Evolution der Schildkröten nach hinten, so dass er innerhalb des Panzers zu liegen kam, der seinerseits die Rippen einschloss. Dafür gibt es jedoch keine Befunde als Anhaltspunkte; in der Embryonalentwicklung heutiger Schildkröten tritt eine solche Bewegung nicht auf (Rieppel 2009, 154).
Die notwendigen Umbauten erforderten nicht nur Veränderungen im Skelettbau, sondern abgestimmte Änderungen anderer Teile des Bauplans. So ist die Muskulatur stark verändert und einige Muskeln haben neue Ansatzstellen (Nagashima et al. 2009). Dass dafür geringere Anpassungen von Entwicklungsabfolgen ausreichen (Joyce 2009, 512), muss bezweifelt werden. So erfordert die Ausbildung neuer Muskelansätze innovative Änderungen, die mehr als bloße Anpassungen beinhalten. Um die Tragfähigkeit evolutiver Modelle beurteilen zu können, müssen die genetischen und entwicklungsbiologischen Details bekannt sein. Wie erwähnt ist hier vieles ungeklärt, weshalb darauf aufbauende evolutionäre Hypothesen nur vage formuliert werden können. Dabei ist immer zu bedenken, dass einzelne Strukturen immer in der Wechselwirkung mit anderen gesehen werden müssen.
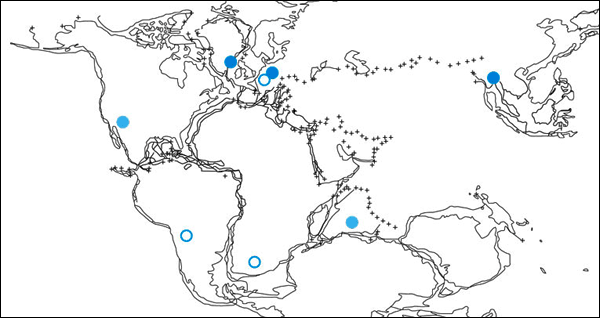 |
| Abb. 4: Paläogeographische Verbreitung der Proganochelyidae (blau), der Australochelyidae (hellblau) und ursprünglicher Halsberger-Gattungen (offene Kreise) aus dem Unterjura. (Nach Karl & Tichy 2000) |
|
Die ältesten Fossilfunde passen zu den konkurrierenden Hypothesen
Zweifellos unterstützen die ältesten Schildkrötenfunde insofern evolutionstheoretische Deutungen, als diese Gattungen (wie Proganochelys) mutmaßliche Primitivmerkmale besitzen, z. B. in der Kopfmorphologie, und dass es unter den älteren fossilen Gattungen Tendenzen hin zum Bau der modernen Formen gibt (Gaffney et al. 1987; Gaffney & Kitching 1994; Rougier et al. 1995; Benton 2007, 245ff.; Pritchard 2008). Und manche Merkmale können als intermediär zwischen Schildkröten und mutmaßlichen Vorläufern gelten. Es sei hier vor allem an die Gattungen Odontochelys und Chinlechelys erinnert. Doch diese Befunde sind nur ein Teil einer komplexen Befundlage. Es sind nämlich folgende Punkte zu berücksichtigen:
- Die Bestimmung „ursprünglich“/„abgeleitet“* ist objektiv allein anhand der jeweiligen Merkmalszustände nicht möglich. Diese Kennzeichnung erfolgt im Rahmen phylogenetischer Hypothesen und kann je nach Datenlage wechseln. Dies wird besonders deutlich an der Wertung des anapsiden Schädels der Schildkröten. Diese Schädelform gilt gewöhnlich als ursprünglich; bei den Schildkröten wird sie seit etwa einer Dekade als abgeleitet gewertet. Viele als Homologien betrachtete Merkmale, die früher eine Verwandtschaft der Schildkröten mit den Pareiasauriern oder Procolophoniden begründeten, müssen nach den neueren molekularen Studien als Konvergenzen gewertet werden.
- Die ältesten Funde sind morphologisch sehr facettenreich mit einer mosaikartigen Merkmalsverteilung und müssen deshalb – in einer evolutions-theoretischen Lesart – sehr verschiedenen Linien zugeordnet werden. Während Odontochelys als wasserlebend angesehen wird, war Chinlechelys sehr wahrscheinlich landlebend. In eine evolutive Reihe passen die beiden Gattungen auch aufgrund ihrer Merkmalsausprägungen nicht; sie müssen evolutionstheoretisch auf verschiedene divergente Linien gestellt werden. Die Gattung Proterochersis (ca. 214 Millionen Jahre; s. o.) besitzt einige abgeleitete Merkmale, ist aber älter als die „primitivere“ Gattung Proganochelys (ca. 206 und ca. 212,5 Millionen Jahre: s. o.). Unter den Schildkröten tauchen also fossil zwar die insgesamt „primitiveren“ Formen zuerst auf, aber sie sind von Beginn der Fossilüberlieferung an in deutlich verschiedene Formen aufgefächert. Die widersprüchliche Merkmalsverteilung unter den frühen Formen (Rougier et al. 1995; vgl. Rieppel & de Braga 1996), kann evolutionstheoretisch nur als Hinweis auf eine konvergente Evolution parallel existierender Linien gewertet werden.
- Die beiden ältesten Gattungen Chinlechelys und Odontochelys begünstigen die beiden konkurrierenden Panzer-Entstehungshypothesen; Chinlechelys passt eher zur Transformationshypothese, während die Merkmale von Odontochelys eher für die Emergenzhypothese sprechen. Dieser Umstand nötigt dazu, beide Gattungen evolutionstheoretisch auf verschiedene Äste zu stellen oder Odontochelys als abgeleitet und das Fehlen des Carapax als sekundär zu interpretieren (vgl. Abschnitt „Neue Fossilfunde“).
Der erstere Fall liefe auf eine zweifach unabhängige Entstehung der Schildkröten hinaus, was evolutionstheoretisch sehr problematisch wäre.
Molekulare Daten und der Fossilbefund
Die zeitliche Stellung der Fossilien scheint im Vergleich zur systematischen Position deutlich zu früh zu sein. Wie im ersten Artikelteil erwähnt ermittelten Mannen & Li (1999) aufgrund molekularer Befunde eine Divergenzzeit für die Trennung der Alligatoren- und Schildkrötenlinie von 151,6 +/– 23,7 Millionen Jahren. Die ältesten Schildkrötenfossilien werden dagegen auf etwa 220 Millionen Jahre datiert. Aus der Obertrias (um 200 Millionen Jahre und älter) ist eine große Fülle fossiler Formen bekannt. Wie ist diese deutliche Inkonsistenz von molekularen Daten und dem Fossilbericht zu deuten?
Eine Untersuchung der Vorderextremitäten der nach phylogenetischen Analysen ursprünglichsten Schildkrötengattungen Proganochelys und Palaeochersis durch Joyce & Gauthier (2004) ergab, dass es sich mit hoher Wahrscheinlichkeit um landlebende Schildkröten handelt. Dagegen weisen phylogenetische Studien wasserlebende Formen als ursprünglich aus. Diese Diskrepanz molekularer und paläontologischer Daten könnte durch Odontochelys (s. o.) gemildert werden, falls diese Gattung primär wasserlebend war.
Einzigartige Merkmalskombinationen und Konvergenzen
Im ersten Artikelteil wurde bereits angesprochen, dass die Schildkröten systematisch mit keiner anderen Amniotengruppe gut verbunden werden können. Es gibt für die diskutierten und weit divergierenden systematischen Positionen der Schildkröten jeweils Argumente; die Schildkröten sind ein besonders schwer zu klassifizierendes Taxon (Wilkinson et al. 1997; vgl. Joyce & Gauthier 2004, 1). Die Schildkröten sind zu allen möglichen Gruppen hin im Bau deutlich abgegrenzt. Dendrogramme weisen ein hohes Maß an Konvergenzen auf (vgl. Rieppel & de Braga 1996, 454; Rougier et al. 1995; Hill 2005). Beispielsweise müssen die gleichen Änderungen des Schultergürtels und der Extremitäten in mehreren Linien von wasserlebenden Schildkröten angenommen werden (Depecker et al. 2006); sehr ähnliche Schädel- und Panzerformen kommen bei den Emydidae, Geoemydidae und Testudinidae vor (Claude et al. 2005). Und einen keulenartigen Schwanz besitzt nicht nur eine der ältesten fossilien Gattungen, Proganochelys, sondern 180 Millionen Jahre später auch die Landschildkröte Meiolania platyceps; und die heute lebende Landschildkröte Chelus fimbriatus hat einen auffallend ähnlichen Bau wie Proganochelys. Viele konvergente Entwicklungen müssen auch in den beiden Hauptgruppen der Cryptodira und Pleurodira angenommen werden (Gaffney 1975, 391, 421f.).
Der Befund weitverbreiteter Konvergenzen ist evolutionstheoretisch insofern problematisch, als vernetzte Merkmalsbeziehungen der verschiedenen Taxa aufgrund von Konvergenzen in einem evolutionstheoretischen Baumschema auf höherem taxonomischem Niveau eher die Ausnahme und nicht die Regel sein sollten. Denn in einem Prozess, der ohne jede Zielorientierung abläuft, ist die zwei- oder mehrmalige Entstehung derselben oder einer sehr ähnlichen Struktur sehr unwahrscheinlich.
Probleme des Umbaus
Bei einer Evolution der Schildkröten von Amnioten-Vorfahren müssten sehr weitgehende „Umbau-maßnahmen“ vorgenommen worden sein. Diese Problematik wurde am Beispiel des Panzers angesprochen und soll hier an einem weiteren Beispiel kurz verdeutlicht werden. Die Rippen erfüllen bei landlebenden Vierbeinern Funktionen in der At-mung und der Fortbewegung: Beide Funktionen können sie bei den Schildkröten nicht (mehr) erfüllen. Durch die Verwachsung der Rippen mit dem Panzer fehlt die Beweglichkeit des Brustkorbs. Die Atmung muss daher durch Bewegung über die Extremitäten unterstützt werden und erfordert ganz neue spezielle Atemtechniken und dafür besondere Muskulatur (genauer beschrieben z. B. unter http://www.monta-n.net/sk/atem.html oder unter http://www.ausgabe.natur-lexikon.com/Schildkroeten.php). Der Übergang zu dieser andersartigen Atemtechnik wäre besonders schwierig gewesen, wenn er in landlebenden Reptilien hätte vollzogen werden müssen, da dort die Möglichkeit der Unterstützung durch den Wasserdruck fehlt. In einem hypothetischen Übergangsstadium gäbe es Konflikte in den funktionalen Rollen der Muskulatur der Atmung und des Bewegungsapparats (Rieppel 2008, 349). Evolutionäre Szenarien der Schildkrötenentstehung, bei denen Details des mutmaßlichen Umbaus nicht bedacht werden, dürften wertlos sein.
Ontogenese als Modell für Phylogenese?
Li et al. (2008) begründen die Interpretation von Odontochelys als Vorstufe zu vollgepanzerten Schildkröten unter anderem damit, dass man diese Gattung mit einem embryonalen Stadium heutiger Schildkröten vergleichen könne. Auch Nagashima et al. (2009, 196) bemerken, dass Odontochelys embryonalen modernen Schildkröten in mancher Hinsicht ähnlich sei und ein Vorfahrenstadium repräsentieren dürfte. Dagegen passen die Merkmale von Chinlechelys nicht zu ontogenetischen Befunden. Bei Chinlechelys sind Rippen und Osteodermen* nur lose verbunden, was evolutionstheoretisch als allmähliche Entstehung des Panzers durch Vergrößerung der Hautknochen und deren spätere Verschmelzung mit Rippen und Wirbeln gedeutet wird. Ontogenetisch entsteht der Panzer aber anders. Die Seiten- und Wirbelsäulenschilde
 |
Der ontogenetische Befund wirft mehr
Fragen auf als er hilft, die Phylogenese
der Schildkröten leichter zu verstehen. |
 |
|
bilden sich ontogenetisch nicht unabhängig von den Rippen und den Wirbeln (Gilbert et al. 2001; Joyce et al. 2009, 511), allerdings sprechen Analysen der Gewebe wiederum für eine getrennte Entstehung der später verschmolzenen Elemente.
Die Ontogenese eignet sich angesichts dieser divergierenden Befunde wie so oft nicht als Wegweiser für die stammesgeschichtliche Rekonstruktion. Joyce et al. (2009, 511) drücken das so aus: „Wir argumentieren daher, dass Daten von heutigen Formen alleine den Prozess, durch den der Schildkrötenpanzer entstand, nicht schlüssig aufklären können.“ Wie im Fall der Dinohand (Ullrich 2008) wirft der ontogenetische Befund mehr Fragen auf als er in die Lage versetzen würde, die Phylogenese der Schildkröten leichter zu verstehen. Denn es geht um einen dramatischen Bauplanwechsel, der tiefgreifend im Vergleich zu den nächsten Verwandten der Schildkröten ist. Während von Befürwortern des Evo-Devo-Ansatzes betont wird, dass die Ontogenese an sog. „constraints“ (Entwicklungszwänge) gebunden sei und diese weitere evolutive Entwicklungsmöglichkeiten kanalisieren, muss hier im Gegenteil angenommen werden, dass bisherige constraints weitgehend aufgehoben werden, da ontogenetisch bereits in einem sehr frühen Stadium die Weichen in Richtung Panzerbildung gestellt werden. Mit dem Begriff der „Emergenz“ (vgl. Emergenzhypothese sensu Rieppel) wird hier ein Ergebnis beschrieben, jedoch keine Erklärung geboten.
Es stellt sich abschließend die Frage nach der Deutung der vorgestellten und diskutierten Befunde in einem nicht-evolutionären Szenario. Dazu sollen einige wenige Hinweise gegeben werden, die als spekulativ gekennzeichnet werden müssen, aber Forschung anregen können.
|
|
Nichtevolutionäre Deutung der frühen Schildkrötenfossilien
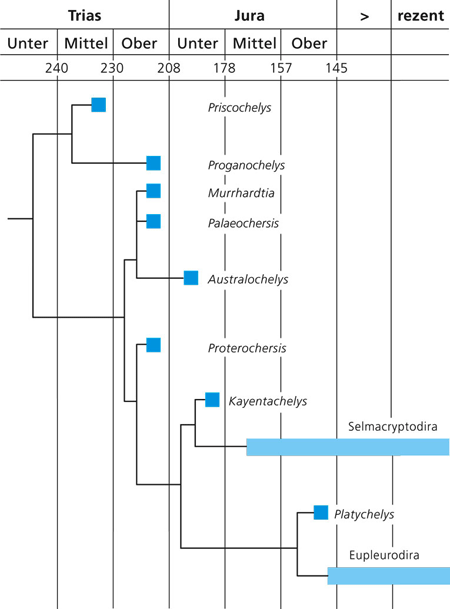 |
| Abb. 5: Auftreten früher Schildkrötengattungen in der Schichtenfolge und vermutete Abstammungszusammenhänge. (Nach Karl & Tichy 2000) |
|
Schnelle große Vielfalt
In der Obertrias sind verschiedene Schildkrötenfossilien bekannt (Abb. 5), die verschiedenen evolutionären Linien zugeordnet werden und sowohl land- als auch wasserlebende Formen beinhalteten. Ähnlich wie bei vielen anderen Tier- und Pflanzengruppen treten verschiedene Linien nebeneinander in enger zeitlicher Folge fossil in Erscheinung, darunter wasser- und landlebende Formen. Neben den als Übergangsformen interpretierbaren Gattungen Odontochelys und Chinlechelys treten fast zeitgleich voll entwickelte Schildkrötengattungen auf. Eine der stratigraphisch ältesten Gattungen, Proterochersis aus dem Unteren Stubensandstein Württembergs ist nach Wild (1998) deutlich „moderner“ als die jüngere Proganochelys, die neben einigen Primitivmerkmalen bereits einen gut ausgebildeten Panzer besitzt (Fraas 1913). Sie gehört wahrscheinlich zu den Halswender-Arten (Pleurodira) (Lucas et al. 2000, 294). Fossilien aus der Untergruppe der Halsberger (Cryptodira) sind ab dem Unterjura bekannt. Nach allen Hypothesen der Schildkrötenverwandtschaft sind die Cryptodira Schwestergruppe* zu den Pleurodira; es sollte also eine Cryptodira-Gattung geben, die so alt ist wie Proterochersis, auch wenn die älteste bisher bekannte Cryptodira-Gattung, Kayentachelys, aus dem Unterjura stammt (Lucas et al. 2000, 294; vgl. Gaffney et al. 1987). Sowohl die Halsberger als auch die Halswender benötigen komplexe, koordinierte Änderungen der Halswirbel und der Muskeln (http://www.earthhistory.org.uk/transitional-fossils/origin-of-turtles/). Lucas et al. (2000) kommen zum Schluss, dass mindestens vier Schildkrötenlinien (Proganochelys, Australochelidae, Pleurodira und Cryptodira) im Revueltium (Stufe der Obertrias) existiert haben müssen (ebenso Lee 1994). Gaffney & Kitching (1994, 57) bezeichnen einige obertriassische Gattungen als lebende Fossilien zu ihrer Zeit.
Diese paläontologisch dokumentierte frühe Vielfalt kann nicht-evolutionär interpretiert werden, wenn eine ökologisch bedingte Fossilisation begründbar ist. Das heißt: Die verschiedenen Schild-krötenformen existierten bereits, traten aber erst dann fossil in Erscheinung, als ihre Lebensbedingungen in so großem Ausmaß verfügbar waren, dass sie sich in genügender Zahl vermehren und ausbreiten konnten. Zuvor lebten sie in geologisch nicht überlieferten Lebensräumen. Diesen Ansatz beschreibt Stephan (2002). Ein solches Szenario kann das Nebeneinander verschiedener Formen möglicherweise besser verständlich machen als ein evolutionäres, wonach ein allmähliches Divergieren eher zu erwarten wäre.
Schnelle weite geographische Verbreitung, kein Ausbreitungszentrum
Das Szenario des ökologisch bedingten Hervortretens aus geologisch zuvor nicht überlieferten Lebensräumen wird durch die schnelle weitweite Verbreitung der ältesten Schildkröten aus der Obertrias unterstützt. Schildkrötenfossilien der Obertrias wurden auf verschiedenen Kontinenten gefunden: in Grönland, Südamerika (Argentinien), Asien (Thailand, China), Europa (Deutschland) und Nord-amerika (Lucas et al. 2000, Karl & Tichy 2000, 66; Joyce et al. 2009, Li et al. 2008, Pritchard 2008, 46; vgl. Abb. 4). Ein paläogeographisches Ausbreitungszentrum zeichnet sich nicht ab (Joyce et al. 2009, 510); evolutionstheoretisch wäre eher ein Ausbreitungszentrum zu erwarten.
|
Das zeitlich gestaffelte Auftreten
verschiedener Gattungen bleibt eine
Herausforderung für nicht-evolutionäre
Erklärungsansätze. |
|
|
Ein ökologisch bedingtes Hervortreten könnte besonders durch geologische Studien überprüft werden: Weisen die Schichten, in denen die Schildkrötenfossilien entdeckt wurden, offenkundige Kennzeichen einer schnellen Bildung auf? Wenn die fossil überlieferten stratigraphisch ältesten Fossilien auf Ausbreitung aus geologisch nicht überlieferten Lebensräumen zurückzuführen sind, lässt sich ein solches geologisches Szenario am besten mit einem kurzen Zeitrahmen von Jahren und Jahrzehnten vereinbaren. Das könnte anhand von Indizien der betreffenden geologischen Schichten überprüft werden.
Eine Herausforderung bleibt auch im ökologischen Ansatz die Erklärung des zeitlich gestaffelten Auftretens verschiedener Gattungen. Warum sind Formen, die am ehesten als Übergangsformen interpretierbar sind, unter den ältesten? Zu prüfen wäre in diesem Zusammenhang, ob die unterschiedlichen Gattungen auf eine Radiation eines oder weniger Grundtypen zurückgeführt werden könnten. Auch diese Frage eröffnet ein weites Forschungsfeld.
|
Literatur
- Benton MJ (2007)
- Paläontologie der Wirbeltiere. München.
- Cebra-Thomas J, Tan F, Sistla S, Estes E, Bender G, Kim C, Riccio P & Gilbert SF (2005)
- How the Turtle Forms its Shell: A Paracrine Hypothesis of Carapace Formation. J. Exp. Zool. 304B, 558-569.
- Claude J, Pritchard P, Tong H, Paradis E & Auffray JC (2005)
- Ecological correlates and evolutionary divergence in the skull of turtles: a geometric morphometric assessment. Syst. Biol. 53, 933-948.
- Depecker M, Berge C, Penin X & Renous S (2006)
- Geometric morphometrics of the shoulder girdle in extant turtles (Chelonii). J. Anat. 208, 35-45.
- Fraas E (1913)
- Proterochersis, eine pleurodire Schildkröte aus dem Keuper. Jh. Ver. vaterländ. Nat.kd. Württ. 69, 13-30.
- Gaffney ES (1975)
- A phylogeny and classification of the higher categories of turtles. Bull. Am. Mus. Nat. Hist. 155, 387-436.
- Gaffney ES, Hutchinson JH, Jenkins FA & Meeker LJ (1987)
- Modern Turtle Origins: The Oldest Known Cryptodire. Science 237, 289-291.
- Gaffney ES & Kitching JW (1994)
- The most ancient African turtle. Nature 369, 55-58.
- Gilbert SF, Loredo GA, Brukmann A & Burke AC (2001)
- Morphogenesis of the turtle shell: the development of a novel structure in tetrapod evolution. Evol. Dev. 3, 47-58.
- Hill RV (2005)
- Integration of Morphological Data Sets for Phylogenetic Analysis of Amniota: The Importance of Integumentary Characters and Increased Taxonomic Sampling. Syst. Biol. 54, 530-547.
- Joyce WG (2007)
- Phylogenetic Relationships of Mesozoic Turtles. Bulletin of the Peabody Museum of Natural History 48, 3-102.
- Joyce WG & Gauthier JA (2004)
- Palaeoecology of Triassic stem turtles sheds new light on turtle origins. Proc. R. Soc. Lond. 271B, 1-5.
- Joyce WG, Lucas SG, Scheyer TG, Heckert AB & Hunt AP (2009)
- A thin-shelled reptile from the Late Triassic of North America and the origin of the turtle shell. Proc. R. Soc. 276B, 507-513.
- Karl HV & Tichy G (2000)
- Murrhardtia staeschei n. gen. n. sp. – eine neue Schildkröte aus der Oberen Trias von Süddeutschland. Joannea Geol. Paläont. 2, 57-72.
- Lee MSY (1994)
- The turtleís long-lost relatives. Nat. Hist. 103, 63-65.
- Li C, Wu XC, Rieppel O, Wang LT & Zhao LJ (2008)
- An ancestral turtle from the Late Triassic of southwestern China. Nature 456, 497-501.
- Lucas SG, Heckert AB & Hunt AP (2000)
- Probable turtle from the Upper Triassic of East-Central New Mexico. N. Jb. Geol. Paläont. Mh. 2000 (5), 287-300.
- Maisch M (2009)
- Trias-Schildkröten aus China und New Mexico. Nat. Rdschau 162, 199-201.
- Mannen H & Li SSL (1999)
- Molecular evidence for a clade of turtles. Mol. Phyl. Evol. 13, 144-148.
- Nagashima H, Uchida K, Yamamoto K, Kuraku S, Usuda R & Kuratani S (2005)
- Turtle„Chicken Chimera: An Experimental Approach to Understanding Evolutionary Innovation in the Turtle. Dev. Dyn. 232, 149-161.
- Nagashima H, Sugahara F, Takechi M, Ericsson R, Kawashima-Ohya Y, Narita Y & Kuratani S (2009)
- Evolution of the Turtle Body Plan by the Folding and Creation of New Muscle Connections. Science 325, 193-196.
- Pritchard PCH (2008)
- Evolution and Structure of the Turtle Shell. In: Wyneken J, Godfrey MH & Bels V (eds) Biology of Turtles. Boca Raton: CRC Press, S. 45-83.
- Reisz RR & Head JJ (2008)
- Palaeontology: Turtle origins out to sea. Nature 456, 450-451.
- Rieppel O (2008)
- The relationships of turtles within amniotes. In: Wyneken J, Godfrey MH & Bels V (eds) Biology of Turtles. Boca Raton: CRC Press, S. 345-353.
- Rieppel O (2009)
- How did the turtle get its shell? Science 325, 154-155.
- Rieppel O & de Braga M (1996)
- Turtles as diapsid reptiles. Nature 384, 453-455.
- Rougier GW, de la Fuente MS & Arcucci AB (1995)
- Late Triassic turtles from South America. Science 268, 855-858.
- Stephan M (2002)
- Der Mensch und die geologische Zeittafel. Holzgerlingen.
- Ullrich H (2008)
- Sind Vogelflügel umgestaltete Dinosaurierhände? Zum Konflikt zwischen fossilen und entwicklungsbiologischen Daten bei der phylogenetischen Herleitung eines Vogelflügels. Stud. Int. J. 15, 18-30.
- Wild R (1998)
- Zwischen Land und Meer. Saurier der Keuperzeit. In: Heizmann EPJ (Hg.) Vom Schwarzwald zum Ries, München, S. 57-66.
- Wilkinson M, Thorley J & Benton MJ (1997)
- Uncertain turtle relationships. Nature 387, 466.
-
|
|  |
