 |
 Studium Integrale Journal
Studium Integrale Journal
15. Jahrgang / Heft 2 - Oktober 2008
Seite 111 - 120
|
|
| |
|
Bereits 1974 wies die Raumsonde Mariner 10 nach, dass der Planet Merkur ein schwaches Magnetfeld besitzt. Merkur und die Erde sind die einzigen der inneren Planeten, die ein Magnetfeld aufweisen, wobei das Magnetfeld von Merkur nur etwa 1% der Stärke desjenigen der Erde besitzt. Es war bislang unklar, wodurch sein Magnetfeld erzeugt wird. Unter anderem wurde spekuliert, dass das jetzige Magnetfeld der Überrest eines früheren Magnetfeldes ist und durch magnetische Strukturen in der Kruste aufrecht erhalten wird, während der Kern mittlerweile abgekühlt und fest ist (Salomon et al. 2008).
Anfang dieses Jahres flog die unbemannte Raumsonde MESSENGER zum ersten Mal an Merkur vorbei. Sie vermaß dabei unter anderem das Magnetfeld (Anderson et al. 2008). Überraschend war, dass die Intensität des Magnetfeldes seit der Entdeckung 1974 praktisch gleich geblieben zu sein scheint. Unterschiedliche geometrische Untersuchungen führen zu dem vorläufigen Ergebnis, dass es genauso wie das Magnetfeld der Erde durch einen magnetischen Dipol erzeugt wird. Die Autoren schließen daraus, dass der äußere Kern des Merkur flüssig und somit für das Magnetfeld verantwortlich sei. Die These, dass das Magnetfeld durch magnetische Strukturen in der Kruste aufrecht erhalten wird, scheint damit durch die Ergebnisse von MESSENGER sehr unplausibel geworden zu sein.
Zusätzlich untersuchte MESSENGER die Magnetosphäre (Slavin et al. 2008), welche durch die Wechselwirkung des Magnetfeldes mit dem Sonnenwind – bestehend aus geladenen Teilchen – erzeugt wird. Sie hat den Messungen zufolge eine ähnliche Form wie der Schweif eines Kometen. Dies ist auf die kurze Distanz Merkurs zur Sonne zurückzuführen, da der Sonnenwind bei dieser Distanz eine hohe Geschwindigkeit hat. Eine detaillierte Untersuchung der Magnetosphäre unterstützt die Hypothese, dass das Magnetfeld durch einen flüssigen äußeren Kern hervorgerufen wird.
Ähnliches trifft auch auf die Untersuchung der sogenannten Exosphäre zu. Die Exosphäre setzt sich aus geladenen Teilchen in der unmittelbaren Umgebung der Oberfläche des Planeten zusammen. Bewegen sich geladene Teilchen durch ein Magnetfeld, so werden diese abgelenkt. Aus dem Verhalten der geladenen Teilchen in der Exosphäre des Merkur können somit Rückschlüsse auf das Magnetfeld gezogen werden. Eine mit Hilfe von MESSENGER durchgeführte Untersuchung der Exosphäre unterstützt ebenfalls die Hypothese, wonach der Merkur einen flüssigen äußeren Kern besitzt (Zurbuchen et al. 2008).
Die Sonde wird noch zwei Mal an Merkur vorbeifliegen und im Jahr 2011 in eine Umlaufbahn um Merkur einschwenken. Dabei sollen unter anderem weitere Messungen des Magnetfeldes, der Magnetosphäre sowie der Exosphäre vorgenommen werden. Sollte dies die bisherigen Ergebnisse bestätigen, so stellt sich die Frage, warum der äußere Kern trotz der geringen Größe des Merkur noch nicht abgekühlt ist, wenn das Sonnensystem tatsächlich so alt ist wie gemeinhin angenommen.
[Salomon SC et al. (2008) Return to Mercury: A Global Perspective on MESSENGER’s first Mercury Flyby. Science 321, 59; Anderson BJ et al. (2008) The Structure of Mercury’s Magnetic Field form MESSENGER’s first Flyby. Science 321, 82; Slavin JA et al. (2008) Mercury’s Magnetosphere after MESSENGER’s First Flyby. Science 321, 85; Zurbuchen TH et al. (2008) MESSENGER Observations of the Composition of Mercury’s Ionized Exosphere and Plasma Environment. Science 321, 90.] ] |
|

|
Die Raumsonde MESSENGER nahm Anfang des Jahres unterschiedliche Bilder der Merkuroberfläche auf. Ein besonderes Augenmerk lag dabei auf der Zahl und der Größe der vorhandenen Einschlagskrater im sogenannten Caloris Becken (Strom et al. 2008). Dabei handelt es sich um ein großes Becken, das vermutlich vulkanischen Ursprungs ist. Aus der Zahl und der Größe der Einschlagskrater innerhalb des Beckens erhofft man sich Rückschlüsse auf das Alter des Beckens. Zunächst wurden die Durchmesser der auf den Aufnahmen erkennbaren Krater bestimmt. Hieraus wurde für jede mögliche Kratergröße die zugehörige Anzahl von Kratern bestimmt. Die Kratergröße sowie die jeweilige Anzahl von Kratern wurden anschließend in einem Diagramm gegeneinander aufgetragen. Diese Methode wurde bereits mehrfach für Altersbestimmungen auf unterschiedlichen Planeten und Monden im Sonnensystem (z.B. Mond und Mars) verwendet (Strom et al. 2005).
 |
| Abb. 1: Der Sveinsdóttir-Krater ist wahrscheinlich durch den Einschlag eines größeren Meteoriten entstanden und besitzt einen Durchmesser von etwa 220 km. Eine etwa 600 km lange Spalte führt mitten durch den Krater. Es wird vermutet, dass diese Spalte durch eine langsame Abkühlung der Merkuroberfläche entstanden ist. (NASA) |
|
Für große Werte der Kratergrößen war in den zugehörigen Diagrammen ein einfacher Zusammenhang in Form einer Potenzfunktion gefunden worden. Das bedeutet, dass die Anzahl Krater der Größe x proportional zu x–n ist, wobei die Potenz n für alle Größen gleich ist. Es wurde davon ausgegangen, dass die zugehörigen Krater mehrheitlich während einer frühen Epoche des Sonnensystems durch die Einschläge einer großen Zahl von Asteroiden aus dem Asteroidengürtel zwischen Mars und Jupiter entstanden sind. Für kleinere Kratergrößen ergab sich ebenfalls eine Potenzfunktion, der Exponent hatte hier jedoch einen anderen Wert. Für die kleinen Krater lautet die Hypothese, dass sie mehrheitlich durch sogenannte sekundäre Einschläge entstanden sind. Dabei handelt es sich um Material, das bei Einschlägen weggeschleudert wird und später erneut auf der Oberfläche auftrifft. Die Zahl der Krater einer bestimmten Größe setzt sich damit aus primären und sekundären Kratern zusammen, wobei unterhalb einer bestimmten Größe die sekundären Krater gegenüber den primären überwiegen. Die Kratergröße, bei der die Zahlen der primären und der sekundären Krater gleich sind, wird dazu verwendet, die Verteilung der sekundären Krater zu charakterisieren.
Ein Vergleich zwischen den früheren Ergebnissen auf Mond und Mars einerseits und den von MESSENGER gelieferten Aufnahmen andererseits führte nun zu einem überraschenden Ergebnis: Für große Kratergrößen stimmte die Verteilungsfunktion für die verschiedenen Himmelskörper überein. Es wäre zu erwarten, dass dies auch für die sekundären Krater zutrifft. Für die kleineren, vermutlich durch sekundäre Einschläge entstandenen Krater wich die Verteilung im Caloris Becken jedoch entgegen der Erwartung von den Verteilungen auf Mond und Mars ab. Der Übergang, ab dem die sekundären Krater gegenüber den primären Kratern überwiegen, lag bei Merkur bei einem Durchmesser von ca. 10 km, während dieser Übergang bei Mars bei etwa 1 km lag. Die Werte der Exponenten waren für die Verteilungen auf Mond, Mars und Merkur zwar gleich, auf dem Merkur war die Dichte der sekundären Krater jedoch größer als auf dem Mond und dem Mars. Die sekundären Krater spielen somit auf dem Merkur eine größere Rolle als auf dem Mars.
Warum diese kleineren Krater auf dem Merkur eine andere Verteilung aufweisen als auf dem Mond und dem Mars ist unklar (Strom et al. 2008). Die Autoren vermuten, dass die Zusammensetzung der Oberfläche von Merkur dazu führt, dass das bei Einschlägen aufgeschleuderte Material eine andere Größe aufweist als auf dem Mond und dem Mars. Sie kommen zu dem Schluss, dass diese Methode zur Altersbestimmung auf Merkur nur „mit Vorsicht“ angewendet werden sollte. In der genannten Arbeit wurden aus der Verteilung der sekundären Krater jedoch keine konkreten Altersangaben abgeleitet. An dieser Stelle sei noch angemerkt, dass die Autoren nicht der Frage nachgehen, ob die Daten auch durch einen anderen funktionalen Zusammenhang als ein Potenzgesetz angemessen beschrieben werden können. Der vorausgesetzte potentielle Zusammenhang geht aus den in der Arbeit dargestellten Graphiken nicht zwingend hervor.
Bereits in einer früheren Untersuchung der Jupitermonde Io und Europa war die Verteilung der sekundären Krater von dem erwarteten Verlauf abgewichen. Eine mögliche Erklärung hierfür wird in Zahnle et al. (2007) und Alvarellos et al. (2007) vorgestellt. Es wird vermutet, dass bei Einschlägen auf Io aufgeschleuderte Gesteinsbrocken später auf Europa eingeschlagen sind. In dem vorgeschlagenen Modell wird zudem der Einfluss der anderen Jupitermonde untersucht. Die Autoren vermuten zudem einen Zusammenhang zwischen Einschlägen auf dem Mars und Sekundärkratern auf den Jupitermonden.
In den genannten Arbeiten wird somit Folgendes deutlich: Die Größenverteilung der Krater kann nicht ohne weiteres zur Altersbestimmung herangezogen werden, wenn nur die Krater auf einem Himmelskörper isoliert betrachtet werden. Zudem ist die Größe des bei den ersten Einschlägen aufgeworfenen Materials nicht bekannt. Diesbezüglich müssen Annahmen vorgenommen werden. Betrachtet man die Diskrepanzen zwischen den Kraterverteilungen auf Io, Europa, dem Mars, dem Mond sowie dem Merkur, so stellt sich die Frage, ob die Kraterstatistiken für sekundäre Krater überhaupt ein zuverlässiges Instrument zur Altersbestimmung darstellen können.
[Strom RG et al. (2008) Mercury Cratering Record Viewed from MESSENGER’s first Flyby. Science 321,79; Strom RG et al. (2005) The Origin of Planetary Impactors in the Inner Solar System. Science 309,1847; Zahnle KJ et al. (2007) Secondary and sesquinary craters on Europa, Icarus 194, 660; Alvarellos JL et al. (2007) Transfer of mass from Io to Europa and beyond due to cometary impacts, Icarus 194, 220.]]
|
|
Arbeiterinnen der Honigbienen (Apis, Apidea) geben Informationen über entdeckte Futterquellen an Schwarmmitglieder weiter. Die meist differenzierte Form der Informationsweitergabe für weit entfernte Futterquellen ist der so genannte Schwänzeltanz. Dabei bewegt sich die tanzende Biene etwa in Form einer Acht und bewegt ihren Körper auf der Strecke zwischen den sich berührenden Kreisen horizontal hin und her („schwänzeln“). Wie Karl von Frisch in umfangreichen Studien eingehend dokumentiert hat, wird durch die Ausrichtung der Schwänzelstrecke die Richtung zur Futterquelle in Bezug zur Sonne und durch die Dauer des Schwänzelns die Entfernung derselben mitgeteilt. Dies stellt die einzige bisher bekannte Form symbolischer Kommunikation unter Wirbellosen dar.
Weltweit sind neun Arten von Honigbienen beschrieben, die sich nach phylogenetischen Studien (auf DNA-Sequenzdaten basierend) seit 6 bis 8 Millionen Jahren unabhängig voneinander entwickelt haben sollen.
In einer Studie haben Su et al. (2008) zunächst dokumentiert, dass Schwärme der asiatischen Honigbiene (Apis cerana cerane, Acc) und der europäischen Honigbiene (Apis mellifera ligustica, Aml) in derselben Umgebung unterschiedlich kommunizieren. Die Autoren gebrauchen dafür die Bezeichnung „unterschiedliche Tanz-Dialekte“ (different dance dialects). Die Dialekte zeigen sich vor allem in signifikant unterschiedlich langen Schwänzeldauern für eine entsprechende Distanz der Futterquelle vom Bienenstock. Diese steigt mit wachsender Entfernung zwischen Bienenstock und Futterquelle an. Für ihre Untersuchungen haben Su und Mitarbeiter Mischkolonien erzeugt. Dazu brachten sie zu einem Volk aus einer Acc-Königin und 5 000 Acc-Arbeiterinnen zwei Rahmen mit gedeckelten Waben von ca. 5 000 Aml-Puppen. Nach ca. 12 Tagen war ein Bienenmischvolk etabliert, das aus je ca. 5 000 Acc- und Aml-Arbeiterinnen bestand. Umgekehrt konnte aus Aml-Königin und Aml-Arbeiterinnen mit Acc-Brut kein Mischvolk etabliert werden, da die Aml-Arbeiterinnen die geschlüpften Acc-Arbeiterinnen innerhalb von 2-3 Tagen töteten und aus dem Stock entfernten. Die Bienenarten behalten auch im Mischschwarm ihre Dialekte bei, d.h. die Dauer des Schwänzelns für eine bestimmte Entfernung unterscheidet sich bei Tänzerinnen im homogenen Schwarm nicht signifikant von derjenigen im Mischvolk.
In verschiedenen Experimenten wurde nun untersucht, inwieweit im Mischschwarm Tänzerinnen der beiden Arten ihre Informationen an Sammlerinnen der jeweils anderen Art weitergeben und diese zum Futtersammeln an der angezeigten Quelle veranlassen konnten. Die Beobachtungen zeigten, dass die Angaben zur Entfernung der Futterquelle von Sammlerinnen von den Tänzerinnen der anderen Art korrekt aufgenommen und umgesetzt werden konnten, obwohl in ihrer eigenen Art die Entfernung mit einer anderen Schwänzeldauer dargestellt wird. Auch wenn gleichzeitig mehrere Futterquellen angeboten wurden, flogen die instruierten Sammlerinnen die angezeigte Quelle signifikant häufiger an. Dabei scheint die asiatische Acc-Honigbiene den Dialekt der europäischen Aml-Honigbiene besser zu verstehen als umgekehrt.
Für die Autoren unterstreichen diese Beobachtungen, dass bei Bienenarten in einem gemischten Schwarm nach lang zurückliegender Trennung nicht nur der Tanz an sich, sondern auch dessen Interpretation in den Bienen hochgradig konserviert sein müssen. Dies eröffnet spannende Fragen zum „sozialen Lernen“ am Studienobjekt Honigbiene. (Klassische Definition für „soziales Lernen“: Lernen aufgrund von Beobachtung oder Interaktion mit einem anderen Individuum, typischerweise derselben Art, oder dessen Produkte.)
Ob die bisherigen Beobachtungen verträglich sind mit der üblichen Ansicht, dass die symbolische Tanzsprache der Bienen aus primitiven Vorformen der Kommunikation entwickelt worden sei, sollte in weiteren Untersuchungen – die angekündigt sind – geprüft werden. Vorerst ist diese Annahme der primitiven Vorformen bestenfalls eine Arbeitshypothese.
[Su S, Cai F, Si A, Zhang S, Tautz J & Chen S (2008) East Learns from West: Asiatic Honeybees Can Understand Dance Language of European Honeybees. PLoS ONE 3(6): e2365. doi:10.1371/journal.pone.0002365]
|
 |
| Abb. 1: Das etwa 8 cm lange Lanzettfischchen Amphioxus. Die Chorda ist ein elastischer Stützstab oberhalb des Darms, der auch in der Embryonalentwicklung der Wirbeltiere als erstes Stützorgan gebildet wird. Das glashelle Lanzettfischchen lebt im Sand der Meeresküsten (unten). (Nach Portmann 1976) |
|
Nach dem Reaktorunfall von Tschernobyl im Jahr 1986 waren viele Experten zunächst überzeugt, dass die ausgetretene radioaktive Strahlung in Europa viel zu gering sei, um Wanzen oder andere Lebewesen zu beeinträchtigen. Cornelia Hesse-Honegger und Peter Wallimann beschreiben und illustrieren jedoch ein schockierendes Ausmaß an Deformationen bei Wanzen aus Schwedens Fallout-Gebieten. Hesse-Honegger sammelte daraufhin mehr als zwei Jahrzehnte lang in verschiedenen Gebieten über 16.000 Individuen westeuropäischer Wanzen, untersuchte sie genau und identifizierte verschiedene Arten von Fehlbildungen, z. B. fehlende Fühlersegmente, verformte Flügel, asymmetrische Leibsegmente, Geschwüre, schwarze Flecken oder eine veränderte Pigmentierung. In manchen Gebieten nahe Kernkraftwerken und Atomaufbereitungsanlagen betrug der Anteil missgebildeter Individuen bis über 30% der untersuchten Wanzen. Dieser Anteil ist 10-30 mal so hoch wie bei Populationen in weitgehend unbelasteten Lebensräumen.
Es zeigte sich, dass nicht die Entfernung von einer Atomanlage entscheidend ist, sondern Windrichtung und Landschaftsform: Gebiete im „Windkanal“ einer Atomanlage sind wesentlich stärker von Missbildungen betroffen als geschützte Stellen. Die Radionuklide werden in den Wirtspflanzen der Wanzen angesammelt. Eine zwar niedrige, aber lang andauernde Strahlendosis kann weit schädlicher sein als eine kurzzeitige hohe Dosis. Besonders gefährlich sind alpha- und beta-Teilchen, da die entsprechenden Nuklide vom Organismus aufgenommen werden, der dadurch von innen durch die Strahlung belastet wird. Wanzen scheinen darauf besonders empfindlich zu reagieren und könnten als sensitive „Bioindikatoren“ für zukünftige Studien dienen.
Hesse-Honegger empfiehlt, die gegenwärtigen Schwellenwerte für radioaktive Immissionen neu zu überdenken. Allerdings sei die biologische Wirkung einer Strahlung sehr schwer zu beziffern (http://www.chembiodiv.ch/highlight.htm).
Die von Hesse-Honegger dokumentierten und hauptsächlich durch radioaktive Strahlung verursachte Schädigungen der Insekten bestätigen bisherige Erfahrungen über Mutationen. Aber auch in dieser gründlichen und umfangreichen Langzeitstudie finden sich keine Hinweise auf wesentliche, sich positiv auf die Vitalität der Tiere auswirkende Veränderungen. Das Ausmaß der Strahlenschäden könnte zudem als Hinweis dafür gesehen werden, dass eine nennenswerte Steigerung der natürlichen Mutationsrate nicht ohne Schaden für die Überlebensfähigkeit der Art ist.
[Hesse-Honegger C & Wallimann P (2008) Malformation of True Bug (Heteroptera): a Phenotype Field Study of the Possible Influence of Artificial Low-Level Radioactivity. Chemistry & Biodiversity 5, 499-539. (doi: 10.1002/cbdv. 200800001)]
|
|
Die Juwelenwespe Ampulex compressa sorgt ganz reizend für ihre Nachkommen. Sie jagt Kakerlaken als lebendigen Speisevorrat. Die Wespe sticht dazu die Kakerlake mit ihrem Stachel. Allerdings injiziert die Wespe ihr Gift genau in den Gehirnnervenknoten (zerebrales Ganglion) und bewirkt dadurch eine lang andauernde Bewegungshemmung. Die Kakerlake ist zwar bei Bewusstsein, jedoch fängt sie nicht von sich aus an zu laufen und reagiert auch nicht auf Reize, die sie normalerweise flüchten lässt. Das Gift der Wespe wirkt auf die Kakerlake aber nicht generell betäubend oder einschläfernd, sondern erhöht allein die Reizschwelle, bevor die Kakerlake anfängt zu laufen. Der Bewegungsgenerator, der die Bewegung der Beine steuert und koordiniert, ist an sich nicht betroffen und so kann die Wespe ihre Beute am Fühler nehmen und wie einen Hund an der Leine in ihr Nest dirigieren. Auch alle anderen Bewegungsauslöser, z. B. Aufrichten nach Umkippen oder ein Sprung bei einer plötzlichen Berührung des Hinterleibes usw. sind vom Gift nicht betroffen. Im Nest angekommen, legt die Wespe ein Ei auf die Kakerlake und versiegelt das Nest. Dort wird die Kakerlake von der schlüpfenden Wespenlarve allmählich aufgefressen.
[Gal R & Libersat F (2008) A parasitoid wasp manipulates the drive for walking of its cockroach prey. Curr. Biol. 18, 877-882.]
|
 |
| Abb. 1: Anna-Kolibri (Calypte anna). © SeanW-fotolia.com |
|
Kolibris stehen nicht im Ruf, besonders sangesfreudige Vögel zu sein. Einige Arten können aber durchaus zwitschern und diese Töne gehen offensichtlich auf ihre Stimme zurück. Bei anderen Tönen, die diese Vögel erzeugen können, war man sich diesbezüglich aber nicht so sicher. Wissenschaftler haben nun herausgefunden, dass das besondere Zirpen des Anna-Kolibris auf dessen vibrierende Schwanzfedern zurückgeht. Dieser Kolibri macht während seiner Brautschau Sturzflüge mit einer Geschwindigkeit von bis zu 80 km/h. Während dessen öffnet er kurz seine Schwanzfeder zu einem breiten Fächer und erzeugt dadurch zirpende Töne von bis zu 60 Millisekunden Dauer. Durch die besondere Bauart der Federn und durch den hindurchfegenden Wind fangen diese an zu vibrieren, ganz ähnlich wie das Holzblättchen einer Klarinette. Auf diese Weise kann der Kolibri lautere Töne hervorbringen, als er es je mit seiner Stimme schaffen würde. Diese außerstimmliche Tonerzeugung beim Anna-Kolibri ist die erste überhaupt bei Vögeln bekannte, aber es ist gut möglich, dass auch andere Kolibri-Arten mit ähnlich geformten Schwanzfedern denselben Mechanismus anwenden. Es war nicht ganz einfach herauszufinden, wie diese Kolibris die Töne erzeugen, aber Hochgeschwindigkeitsaufnahmen brachten die Forscher auf die richtige Spur, dort konnte man das Öffnen und Schließen des Schwanzes nachverfolgen. Nachdem man einigen Kolibris die Schwanzfedern ein wenig gestutzt hatte, waren sie nicht mehr imstande, das Zirpen zu erzeugen. Nach fünf Wochen sind die Schwanzfedern erneuert, die Vögel erleiden also keinen bleibenden Schaden. Des Weiteren konnten die Forscher im Windkanal Töne mit den Schwanzfedern der Kolibris erzeugen, wenn die Windgeschwindigkeit besagte 80 km/h erreichte. Die Forscher möchten nun weitere Kolibris aus derselben Gruppe untersuchen und vermuten nun auch, dass es möglicherweise bei Schnepfen eine ähnliche Lauterzeugung geben könnte. Sturzflug und Schwanzmorphologie seien ko-evolviert, um eine Vielfalt an mechanisch erzeugten Lauten hervorzubringen. Sollte es sich tatsächlich erweisen, dass Schnepfen ebenfalls fähig sind, solche Laute zu erzeugen, wäre das auch ein interessanter Fall von konvergenter Evolution – oder auch nicht.
[Williams N (2008) Tail spins. Curr. Biol. 18, R143-144]
|
|
Das Eindringen von neuen Arten in bisher nicht besiedelte Lebensräume bietet einzigartige Gelegenheiten, um die Anpassung dieser Arten an neue Lebensbedingungen zu studieren. Facon und Mitarbeiter (2008) berichten über die Invasion einer Süßwasser-Schnecke (Melanoides tuberculata) auf der Insel Martinique. In einer einzigartigen Studie wurden über Jahre alle Süßgewässer der Insel beprobt und insgesamt 17.000 Schnecken gesammelt. Die Schnecken wurden genetisch und morphologisch (den Körperbau betreffend) untersucht und klassifiziert. Es zeigte sich, dass es sieben verschiedene Populationen dieser Schnecken in Martinique gibt. Fünf davon sind auf jeweils ein Einwanderungsereignis zurückzuführen, während zwei Rassen auf Martinique durch Kreuzung der bereits eingewanderten Rassen entstanden sind. Diese neuen Kreuzungen zeigen nun jeweils Eigenschaften, die es so bei ihren Elternarten nicht gegeben hat, z. B. weniger, dafür aber größere Nachkommen. Die Autoren dieser Studie berichten nun, dass die Hauptursache für die genetische Variabilität einer solchen Invasion die verschiedenen Einwanderungsereignisse seien. Die stattgefundenen Kreuzungen hingegen sollen zeigen, dass solche Invasionsereignisse „kreativ“ sind, denn sie bringen Genotypen zusammen, die bisher nicht die Möglichkeit hatten, miteinander zu hybridisieren. Die Nachkommen sind sprunghaft („saltatory“) verschieden von den Elternarten. Dadurch seien Invasionen auch immer Hotspots, die evolutionäre und ökologische Neuheiten hervorbringen. Es bleibt aber festzuhalten, dass trotz aller „Neuigkeiten“ sich die Evolution dieser Süßwasserschnecken natürlich im Bereich der Mikroevolution bewegt. Die Autoren halten beispielsweise selber fest, dass andere Rassen dieser Schnecken extremere morphologische Werte aufweisen als die neuen durch Kreuzung entstandenen. Diese neuen Kreuzungsrassen bilden also nur einen kleinen Ausschnitt der weltweiten Variabilität dieser Schnecken ab. Die Vermischung oder Selektion bereits vorhandener Merkmale invasiver Arten als „evolutionären Hotspot“ zu bezeichnen, scheint daher doch etwas hoch gegriffen.
[Facon B, Pointier J-P, Jarne P, Sarda V & David P (2008) High genetic variance in life-history strategies within invasive populations by way of multiple introductions. Curr. Biol. 18, 363-367.]
|
|
Einem unscheinbaren wurmartigen glashellen Geschöpf, das im Sand der Meeresküsten lebt, wird eine besondere Stellung im System der Lebewesen zugebilligt: Das etwa 8 cm lange Lanzettfischchen (Branchiostoma lanceolatum; im Englischen nach dem alten Gattungsnamen Amphioxus genannt; Abb. 1) wird an die Basis der Chordatiere gestellt, zu denen auch der Mensch gehört, seit man entdeckt hatte, dass es eine Chorda dorsalis als elastischen Stützstab oberhalb des Darms besitzt (s. Abb. 1). Eine Chorda wird auch in der Embryonalentwicklung der Wirbeltiere als erstes Stützorgan gebildet und im Laufe der Ontogenese durch die Wirbelsäule weitgehend ersetzt. Eine direkte Ableitung der Wirbeltiere vom Lanzettfischchen ist jedoch nicht möglich, da diese Form dafür zu spezialisiert ist. Das Lanzettfischchen gehört zu den Schädellosen (Acrania oder Cephalochordata), die zusammen mit den Wirbeltieren (Vertebrata) und den Manteltieren (Tunicata oder Urochordata) zu den Chordatieren gehören.
 |
| Abb. 1: Das etwa 8 cm lange Lanzettfischchen Amphioxus. Die Chorda ist ein elastischer Stützstab oberhalb des Darms, der auch in der Embryonalentwicklung der Wirbeltiere als erstes Stützorgan gebildet wird. Das glashelle Lanzettfischchen lebt im Sand der Meeresküsten (unten). (Nach Portmann 1976) |
|
Nun hat eine internationale Arbeitsgruppe unter der Leitung von Nicholas Putnam vom Joint Genome Institute des US-amerikanischen Energieministeriums in Walnut Creek und Linda Holland von der University of California in San Diego das Genom (Erbgut) des Lanzettfischchens entschlüsselt. Einige Ergebnisse sollen kurz vorgestellt und diskutiert werden.
Komplexität des Erbguts. Einmal mehr erweist sich das Erbgut eines vergleichsweise einfach gebauten Organismus als ziemlich komplex. Das 520 Millionen Basenpaare lange Genom enthält mit 21 900 proteincodierenden Abschnitten nicht viel weniger Gene als der Mensch. Auch in den nicht-codierenden Bereichen gibt es zahlreiche Übereinstimmungen. Angesichts der langen evolutionären Trennungszeit von Mensch und Lanzettfischchen findet Gee (2008) in einem Kommentar diese Übereinstimung „bemerkenswert“. Die großen genetischen Übereinstimmungen machen deutlich, dass die individuelle Entwicklung und die hypothetische Evolution nicht alleine über die Gene verstanden und abgleitet werden können. Vielmehr sollen bei den Wirbeltieren alte Gene neu kombiniert und ihre Funktion und Steuerung verändert worden sein. Wie das funktionieren kann, ist einer der Gegenstände der neueren Forschungsrichtung „EvoDevo“, die die Entstehung von Neuem in der Evolution vor allem in Prozessen der Ontogenese (Individualentwicklung) sucht (vgl. den Artikel über „Evo-Devo“ in dieser Ausgabe).
Die Daten des Gensatzes von Amphioxus legen im Vergleich mit denen der Wirbeltiere nahe, dass sich das Ur-Genom der Wirbeltiere sehr früh, nämlich vor der Trennung der Knorpel- und Knochenfische, zweimal komplett verdoppelt hat. Gee (2008) spricht von einem „genomischen Sturm“. Die dafür erforderlichen Mechanismen liegen im Dunkeln. Wie die Verdopplungen des Erbguts mit dem Neuerwerb von Organen bei den Wirbeltieren verbunden sein könnte, ist ebenso unbekannt. Damit der verdoppelte Gensatz auch für die Ausbildung neu genutzt werden kann, müsste eine umfangreiche Reprogrammierung erfolgen. Mechanismen dafür sind ebenfalls nicht bekannt.
Konflikt Morphologie-Moleküle. Ebenfalls nicht neu ist die Feststellung, dass die verfügbaren Daten zu den Ähnlichkeiten auf genetischer Ebene nicht mit den Merkmalsübereinstimmungen auf anatomischer Ebene übereinstimmen. Die lange Zeit favorisierte Ableitung der Chordatiere von Larven sessiler (festsitzender) Manteltiere ist genetisch nicht nachvollziehbar und wird daher aufgegeben (Holland et al. 2008, 9). Denn die Genomanalyse ergab, dass sich – evolutionstheoretisch betrachtet – die Manteltiere später von den Wirbeltieren getrennt haben müssen als Branchiostoma. Als wahrscheinlicher wird jetzt angesehen, dass der hypothetische gemeinsame Vorläufer ein frei lebendes, zweiseitig-symmetrisches wurmartiges Tier war (Holland et al. 2008, 9).
[Gee H (2008) The amphioxus unleashed. Nature 453, 999-1000; Putnam MH, Butts T et al. (2008) The amphioxus genome and the evolution of the chordate karyotype. Nature 453, 1064-1071; Holland LZ, Albalat R et al. (2008) The amphioxus genome illuminates vertebrate origins and cephalochordate biology. Genome Research, doi 10.1101/ gr.073676.107]
|
 |
| Abb. 1: Rekonstruierter Schädel von Ventastega curonica. (Nach Ahlberg et al. 1994) |
|
In den letzten Jahrzehnten wurde eine ganze Reihe von Fossilien des Oberdevons entdeckt, die in den Übergangsbereich Fische – Vierbeiner (Tetrapoden) gestellt werden. Die neuen Funde führten zu einem Paradigmenwechsel bezüglich der Evolution der Vierbeiner-Extremität: Der Übergang von Flossen zu Beinen soll demnach im Wasser entstanden sein und nicht erst im Zuge der Eroberung des Landes. Wir hatten in Studium Integrale Journal in einer Artikelserie die Gründe für dieses Umdenken dargestellt (Junker 2004). Die mittlerweile bekannte Formenvielfalt lässt sich in mancher Hinsicht evolutionär im Sinne stammesgeschichtlicher Abfolgen deuten, da unterschiedliche Merkmalsmosaike von Fisch- und Vierbeinermerkmalen vorkommen, die teilweise als evolutionäre Abfolgen interpretierbar sind. Andererseits werden die wasserlebenden oberdevonischen Vierbeiner als evolutive Sackgasse angesehen, und die Formenvielfalt zeigt auch widersprüchliche Merkmalskombinationen (Konvergenzen, s. u.). Alternativ zu einer evolutionären Deutung der Oberdevon-Tetrapoden kommt auch eine ökologische Deutung in Frage. Die oberdevonischen Formen waren offenbar an ein Leben als im Uferbereich lauernde Räuber angepasst. Ihre besonderen Merkmalskombinationen sind vermutlich von daher zu verstehen.
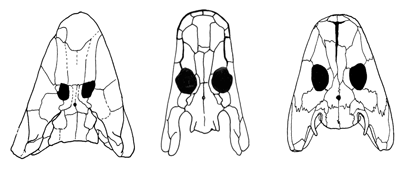 |
| Abb. 2: Schädel von Tiktaalik, Ventastega und Acanthostega von oben. (Nach Ahlberg et al. 2008) |
|
Eine dieser Arten ist Ventastega curonica aus dem oberen Oberdevon Lettlands (Abb. 1). Sie ist schon seit 1994 bekannt und beschrieben (Ahlberg et al. 1994, Clack 2002, 130). Ahlberg und sein Team von der Universität Uppsala veröffentlichten nun weiteres Fundmaterial. Gefunden wurden Schädelteile, Teile des Schultergürtels und ein Teil des Pelvis (Schambein). In vielen Merkmalen (Schädelform, Schultergürtel) ähnelt Ventastega der achtzehigen, wasserlebenden Gattung Acanthostega (vgl. Abb. 2 und 3), andere Merkmale (z. B. die Bezahnung, Teile des Schädels) tendieren mehr zu fischartigen Formen wie Panderichthys. Daher kann Ventastega morphologisch zwischen fischartige Gattungen wie Tiktaalik (vgl. Abb. 2) und wasserlebenden Vierbeinern gestellt werden, wobei insgesamt eine deutlich größere Nähe zu Acanthostega gegeben ist.
 |
| Abb. 3: Rekonstruktion von Acanthostega als schwimmender Vierbeiner (© Richard Hammond) |
|
Trotz vieler fehlender Skeletteile rekonstruieren Ahlberg et al. (2008) bei Ventastega eine Körperform, die Acanthostega stark ähnelt (Abb. 4; ca. 1 m lang). Diese Position ergibt auch eine phylogenetische Analyse (Ahlberg et al. 2008); allerdings treten zahlreiche Konvergenzen (unabhängig entstandene Merkmalsübereinstimmungen) auf, sodass die evolutive Analyse wenig konsistent ist (niedriger Konsistenzindex). Dass es sich bei Ventastega um einen Vierbeiner handelt, wird indirekt anhand der Merkmale von Schulter- und Beckengürtel erschlossen; es sind allerdings keine Teile der Extremitäten entdeckt worden.
 |
| Abb. 4: Rekonstruktion von Ventastega mit den fossil nachgewiesenen Skeletteilen. Maßstab: 10 cm (Nach Ahlberg et al. 2008) |
|
Ahlberg et al. (2008) bestätigen ihre frühere Einschätzung (Ahlberg et al. 1994, 306), dass Ventastega einer der primitivsten Tetrapoden sei. Dies steht allerdings im Gegensatz zur stratigraphischen Stellung, da Ventastega ausgerechnet zu den jüngsten oberdevonischen Tetrapoden gehört. Diese Vierfüßer bilden insgesamt eine ausgeprägt paraphyletische Gruppe (Ahlberg 1995, 423f.), d. h. die Formenvielfalt lässt sich nicht in ein widerspruchsfreies Stammbaumschema bringen; viele Gattungen gelten daher als totes Ende eines Stammbaumastes. Ahlberg et al. (2008, 1203) schätzen auch die Gemeinsamkeiten von Ventastega und Acanthostega als paraphyletisch ein.
Der Übergangsstatus von Ventastega gilt also nicht für alle Einzelmerkmale. So zeigt der Vergleich des Schädeldachs mit den „benachbarten“ Gattungen Tiktaalik und Acanthostega (Abb. 2) eine deutliche Nähe zwischen Ventastega und Acanthostega. Insgesamt ist der Schädel aber nicht intermediär. Ahlberg et al. (2008) weisen am Schluss ihres Artikels darauf hin, dass es keine einfache Linie von Tiktaalik über Ventastega zu Acanthostega gibt. Die Autoren schreiben: „Mindestens zeigt sich die Anwesenheit einer beträchtlichen Diversität von Formen unter den frühesten Tetrapoden.“
[Ahlberg PE (1995) Elginerpeton pancheni and the Earliest Tetrapod Clade. Nature 373, 420-425; Ahlberg PE, Luksevics E & Lebedev OA (1994) The first tetrapod finds from the Devonian (Upper Famennian) of Latvia. Philos Trans. R. Soc. Lond. B 343, 303-328; Ahlberg PE, Clack JA, Luksevics E, Blom H & Zupins I (2008) Ventastega curonica and the origin of tetrapod morphology. Nature 453, 1199-204; Clack JA (2002) Gaining Ground. The origin and evolution of Tetrapods. Bloomington and Indianapolis: Indiana University Press; Junker R (2004) Vom Fisch zum Vierbeiner – eine neue Sicht zu einem berühmten Übergang. Teil 1: Überblick und tetrapodenartige Fische des Oberdevons. Stud. Int. J. 11, 3-10.]
|
|
Eine der ausgeprägtesten Diskontinuitäten in der Fossilüberlieferung findet sich am Übergang vom Präkambrium zum Kambrium. In kambrischen Sedimenten tritt eine hochdifferenzierte Tierwelt so plötzlich und vielfältig auf, dass von der „kambrischen Explosion“ (Valentine 2004) oder vom „Urknall der Paläontologie“ gesprochen wird. Lebewesen aus allen bekannten Tierstämmen, die Hartteile besitzen, sind im Kambrium (meist bereits im Unterkambrium) vertreten. Dazu gehören z. B. Schwämme, Hohltiere, Ringelwürmer, Armfüßer, Gliederfüßer, Weichtiere, Stachelhäuter und auch Chordatiere (darunter sehr wahrscheinlich auch kieferlose Fische als erste Wirbeltiere). Diese Tierstämme sind zudem von Beginn ihres fossilen Nachweises in der Regel in verschiedene, deutlich abgrenzbare Untergruppen aufgespalten und geographisch meist weit verbreitet.
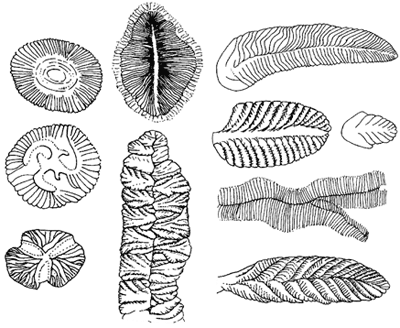 |
| Abb. 1: Die Ediacara-Organismen sind rätselhaft konstruiert: Seilacher sieht in ihnen spezialisierte, einzellige Mikrobenhüllen, „lebende Luftmatratzen“ (mit Protoplasma gefüllte Pneus), ohne innere Organe. Sie sind als Mikrobenfilme auf sandigem Sediment festgeklebt und führten ein uns fremdartig erscheinendes Leben unter weitgehend unbekannten Umweltbedingungen. (Aus Junker & Scherer 2006; nach Gould) |
|
In den unter dem Kambrium abgelagerten Schichten des Präkambriums wurden dagegen vergleichsweise wenige Vielzeller gefunden. Diese werden meist als Ediacara-Fauna bezeichnet (benannt nach dem ersten Fundort, den Ediacara-Bergen in Australien; vgl. Abb. 1). Dabei handelt es sich um eigenartige, hartkörperlose Organismen. Der Paläontologe Seilacher sieht in ihnen spezialisierte, einzellige Mikrobenhüllen, „lebende Luftmatratzen“ (mit Protoplasma gefüllte Pneus), ohne innere Organe; die Körperhüllen sind in sehr verschiedener Weise mit Lamellen abgesteppt. Sie waren teilweise offenbar als Mikrobenfilme auf sandigem Sediment festgeklebt. Es handelt sich um eine isolierte Gruppe, die anderen bekannten Fossilgruppen nicht zugeordnet werden kann; sie verschwindet im Laufe des Kambriums.
Ediacara-Fossilien wurden mittlerweile an über 30 Lokalitäten auf mehreren Kontinenten gefunden und bislang sind mehr als 270 Arten beschrieben worden. Sie werden im System der Historischen Geologie in die Zeitspanne zwischen 575-542 Millionen Jahre datiert und repräsentieren die ältesten bekannten komplexen makroskopischen Lebensformen (Shen et al. 2008, 81). Die Formenvielfalt der ältesten unter ihnen – die auf 575-565 Millionen Jahre datierte Avalon-Gruppe von Neufundland – wurde kürzlich genauer analysiert. Es zeigte sich, dass die bislang bekannte Formenvielfalt schon zu Beginn der Fossilüberlieferung der Ediacara-Fossilien voll ausgeprägt war. Shen et al. (2008) verwenden daher folgerichtig den Begriff „Avalon Explosion“.
Trotz des plötzlichen Auftretens der Formenvielfalt betrachten die Autoren eine längere, undokumentierte evolutive Vorgeschichte als unwahrscheinlich. Das schlagartige, „explosive“ Auftreten ist damit wiederholt dokumentiert und die Frage nach den Mechanismen entsprechend drängend. Die Autoren sehen deutliche Parallelen zur kambrischen Explosion und vermuten in beiden Fällen ähnliche Mechanismen, über die allerdings nur spekuliert werden kann. Als Triebkräfte der plötzlichen Evolution werden z. B. ökologische oder entwicklungsgenetische Gründe (Entwicklung von Regulationssystemen) diskutiert; dabei handelt es sich aber bestenfalls um notwendige Voraussetzungen für größere Veränderungen. Tatsächlich sind solche Explosionen gemessen am bekannten Wissen über evolutionäre Variationsmechanismen überraschend und Mechanismen dafür sind unbekannt. Unter www.sciencedaily.com/... wird M. Kowalewski, einer der Autoren, zitiert (eigene Übersetzung): „Beschleunigte Raten dürften die frühe Evolution vieler Organismengruppen charakterisieren.“ Beschleunigte Evolution ist aber nur im mikroevolutiven Rahmen teilweise verstanden; die Entstehung neuer Baupläne und gar deren beschleunigtes Auftreten sind dagegen Spekulation.
Die Ediacara-Fossilien werden meist als blinde Enden des hypothetischen evolutionären Stammbaums betrachtet; Shen et al. sprechen von einem „gescheiterten Experiment“.
Wie zahlreiche andere Befunde ist auch die Avalon-Explosion im evolutionstheoretischen Rahmen unerwartet, was jedoch weder hier noch sonst zu einem Zweifel am Evolutionsparadigma führt. Der Philosoph H.-D. Mutschler referiert W. Stegmüller, der sogar als analytischer Wissenschaftstheoretiker und Physikalist die Auffassung vertritt, „die biologische Evolutionstheorie sei insofern keine wirklich wissenschaftliche Theorie, da sie oft nur Rekonstruktionen biete, die sich nicht in Vorhersagen verwandeln ließen“ (Mutschler 2002, 159; vgl. 26). Als möglichen Grund für diese Unbeirrbarkeit der evolutionsorientierten Biologie vermutet Mutschler (2003): „Vielleicht ist diese Wissenschaft viel stärker von weltanschaulichen a priori-Entscheidungen durchdrungen, als den entsprechenden Wissenschaftlern bewusst ist.“
[Shen B, Dong L, Xiao S & Kowalewski M (2008) The Avalon Explosion: Evolution of Ediacara Morphospace. Science 319, 81-84; Mutschler H-D (2002) Naturphilosophie. Kohlhammer-TB 396. Stuttgart; Mutscher H-D (2003) Gibt es Finalität in der Natur? In: Kummer C (Hg) Die andere Seite der Biologie. München; Seilacher A (1992) Vendobionta als Alternative zu Vielzellern. Mitt. Hamb. zool. Mus. Inst. 89, Erg.bd.1, 9-20; Valentine (2004) On the origin of phyla. Chicago – London.]
|
|
Befunde aus der Erforschung der Antarktis haben in der jüngsten Zeit immer wieder überraschende Ergebnisse geliefert. Jetzt berichten Mark Williams und Kollegen von Fossilien aus einer Konservat-Lagerstätte, die aus einem See auf dem 77. südlichen Breitengrad stammen. Sie beschreiben einen Muschelkrebs (aus der Klasse der Ostracoda) aus dem Mittleren Miozän, der in Goethit, einem Eisenmineral (FeOOH) erhalten ist, nachdem er ursprünglich pyritisiert war (Pyrit: Eisensulfid, FeS2). Im Miozän (vor ca. 14 Millionen radiometrischen Jahren) war die geographische Lage der Antarktis nach gängigen Vorstellungen bereits ähnlich der heutigen. Wenn damals aber ein Ostracode am Boden des antarktischen Sees den Schlamm kriechend durchwühlen konnte, dann müssen die dort die Temperaturen deutlich wärmer gewesen sein als heute, da die heute bekannten Muschelkrebse unter wärmeren Bedingungen leben.
Nach gängigen Vorstellungen und Theorien hat sich der Eisschild der Antarktis jedoch bereits vor 34 Millionen Jahren zu bilden begonnen. Der neue Fund stellt vor Herausforderungen bezüglich der Prozesse und Geschwindigkeiten der Abkühlung der ökologischen Veränderungen in der Antarktis.
[Williams M, Siveter DJ, Ashworth AC, Wilby PR, Horn DJ, Lewis AR & Marchant DR (2008) Exceptionally preserved lacustrine ostracods from the Middle Miocene of Antarctica: implications for high-latitude paleoenvironment at 77° south. Proc. Roy. Soc. B, Online: DOI10.1098/rspb.2008. 0396]
|
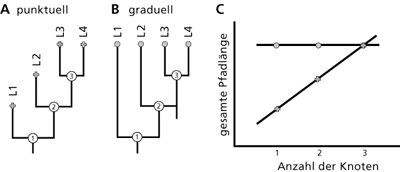 |
| Abb. 1: Punktuelle und graduelle Evolution in Sprachstammbäumen. Punktuelle Evolution (A) postuliert, dass lexikalische Veränderungen sprunghaft bei Auftrennung einer Muttersprache in 2 Tochtersprachen geschehen, so dass die gesamte „Pfadlänge“ und damit das Ausmaß der lexikalischen Veränderungen um so länger ist, je mehr solcher Sprachtrennungsereignisse („Knoten“) in der Geschichte einer Sprache auftraten (ansteigende Linie in C). Graduelle Evolution (B) postuliert, dass es keine Beziehung zwischen der Anzahl der Sprachauftrennungen in der Geschichte einer Sprache und dem Ausmaß der lexikalischen Veränderung gibt (gerade Linie in C ). Nach Atkinson et al. (2008), online support. |
|
Nicht nur biologische, sondern auch kulturelle menschliche Merkmale wie die Sprache sollen durch Evolution entstanden sein (Hauser et al. 2002). In einer kürzlich veröffentlichten Studie wollen Atkinson et al. (2008) entscheiden, ob die Evolution der Sprache(n) vornehmlich graduell oder punktuell erfolgt, d.h. ob sich Sprachen im wesentlichen unabhängig von äußeren Ereignissen mit der Zeit allmählich verändern oder ob Sprachen bei entsprechenden Ereignissen („Knoten“) sich rasch verändern, um anschließend über längere Zeiträume nur wenig zu evolvieren (vgl. Abb. 1). Ein solches äußeres, „punktuelles“ Ereignis könnte z. B. die Kolonisierung eines bisher unbewohnten Gebietes sein, wobei die Gründerpopulation den Kontakt mit ihrer Ursprungspopulation verliert, etwa bei der Neubesiedelung einer Insel. Ein anderes Beispiel wäre die Einwanderung eines anderssprachigen Volkes in ein bereits besiedeltes Gebiet mit anschließender Vermischung beider Populationen.
Atkinson et al. verwenden zur Entscheidung dieser Frage dieselbe Methodik (Webster et al. 2003), die auch in der Molekularbiologie zum Korrelieren der Raten der genetischen Evolution und der Entstehung neuer Arten verwendet wird. Sie untersuchten das Vokabular von 490 Sprachen aus drei großen Sprachfamilien; ein Drittel der Menschheit spricht eine dieser Sprachen: 95 afrikanische Bantusprachen, 65 indogermanische Sprachen und 330 pazifische austronesische Sprachen. Für jede Sprachfamilie griffen sie auf ein „Basisvokabular“ zurück, das nur sehr selten aus einer anderen Sprache übernommen wird. Dieses Basisvokabular umfasste 100 Begriffe in den Bantusprachen, 200 in den indogermanischen Sprachen und 210 in den austronesischen. (Detaillierte Beschreibung der Sprachen und verwendeten Begriffe im Online-Support der Publikation von Atkinson et al.)
Es ergab sich in allen untersuchten Sprachfamilien, dass die lexikalische Veränderung mit der Anzahl der Trennungsereignisse signifikant zunahm (Abb. 2).
 |
| Abb. 2: Pfadlänge aufgetragen gegen die Anzahl der Trennungsereignisse oder „Knoten“: Bantusprachen hellgrau, indogermanische Sprachen dunkelblau, austronesische Sprachen hellblau und deren Untergruppe polynesische Sprachen schwarz. Die eingezeichneten gemittelten und statistisch korrigierten Graden zeigen durch ihre positive Steigung, dass punktuelle und nicht graduelle Evolution bei Sprachen entscheidend ist. Nach Atkinson et al. (2008) |
|
Statistische Verfahren ergaben weiter, dass etwa 31% des Vokabulars der Bantusprachen während der Trennungen neu entstand, 21% bei den indogermanischen und 9,5% bei den austronesischen Sprachen. In deren Untergruppe der polynesischen Sprachen – die typischerweise von nur kleinen, voneinander isolierten auf Inseln im Pazifik lebenden Menschengruppen gesprochen werden – war sogar 33% des Vokabulars bei Trennungsereignissen entstanden Atkinson et al. folgern, dass in der Sprachevolution lange Perioden relativer Konstanz unterbrochen werden durch rasche Veränderung. Sie vermuten, dass dasselbe für die Morphologie und die Syntax gilt. Nach Liebi (2003) zeigen jedoch alle bekannten Sprachen während der gesamten verfügbaren Beobachtungszeit einen ständigen Abbau von morphologischen Strukturen („Formenlehre“), die durch Veränderungen der Syntax zum Teil kompensiert werden. Dieser Befund, der einer von mehreren von Liebi dargelegten Belegen gegen eine Evolution der menschlichen Sprache aus Tierlauten über primitive Vorsprachen bis zu den bekannten Hochsprachen ist, wird von Atkinson et al. nicht berücksichtigt, seine ganze Arbeit suggeriert indirekt, dass die Evolution der Sprache grundsätzlich fachlich unbestritten sei.
Es ist bemerkenswert, dass eine der beiden führenden wissenschaftlichen Fachzeitschriften ihre schon vor 122 Jahren publizierten Richtlinien bezüglich korrekter Methodik zur Sprachevolution nicht beherzigt: „Die gegenwärtige fortgeschrittene Kenntnis unserer Kenntnis der Sprache reflektiert (...) die Vorteile der modernen Forschungsmethodik. Ein herausragendes Kennzeichen dieser Methode ist, einen weiten Übersichtsstandpunkt einzunehmen, von dem aus fast alle relevanten Fakten Interesse und Bedeutung gewinnen: Die moderne Forschungsmethodik sagt nicht eng und pedantisch, dies und das ist mein Arbeitsgebiet und was außerhalb steht, betrifft mich nicht“ (Jastrow 1886).
[Atkinson QD, Meade A, Venditti C, Greenhill SJ & Pagel M (2008) Languages evolve in punctional bursts, Science 319, 588; Hauser MD, Chomsky N & Fitch WT (2002) The faculty of language: what is it, who has it, and how did it evolve? Science 298, 1569-1579; Jastrow J (1886) The evolution of language, Science 7, 555-557; Liebi R (2003) Herkunft und Entwicklung der Sprachen. Linguistik contra Evolution. Holzgerlingen; Webster AJ, Payne RJH & Pagel M (2003), Molecular phylogenies link rates of evolution and speciation, Science 301, 478.]
|
|
Letzte Änderung:
28.01.2009 •
|
 |
|
