|
|
| |
|
Eine Supernova ist eine gewaltige Sternexplosion, bei welcher eine Schockwelle aus stellarem Material (Material von Sternen) in den Weltraum geschleudert wird. Trifft diese Schockwelle auf das interstellare Medium, wird dieses stark erhitzt und emittiert Röntgenstrahlen (d. h. es werden Röntgenstrahlen erzeugt). Dabei bilden sich auch starke Magnetfelder, deren Entstehung jedoch noch nicht in allen Details verstanden ist. Elektronen, die beim Schock beschleunigt werden, senden in diesem Magnetfeld Synchrotronstrahlung aus, die durch das ganze Spektrum von Radiowellen bis Röntgenstrahlung beobachtet werden kann.
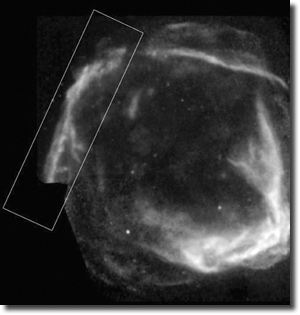 |
| Abb.1: Aufnahmen der Supernova-Überreste RCW 86 im Röntgenbereich mit den Satelliten XMM-Newton und Chandra. Gut sichtbar ist die expandierende Schale des ehemals explodierten Sterns. (Chandra: NASA/CXC/Univ. of Utrecht/J. Vink et al., XMM-Newton: ESA/Univ. of Utrecht; http://chandra.harvard.edu/photo/2006/rcw86/) |
|
Bereits in den 1960er Jahren wurde vermutet, dass die Supernovaüberreste mit der Bezeichnung RCW 86 zur Supernova AD 185 gehören könnten, die chinesische Astronomen im Jahr 185 n. Chr. aufgezeichnet hatten. An diesem Zusammenhang wurden jedoch seit Ende der 1980er Jahre zunehmend Zweifel geäußert. Der Grund dafür war, dass RCW 86 einerseits offenbar zu weit entfernt sei und andererseits viel älter als das historische Alter von AD 185 (z.B. Rosado et al. 1996). Aufgrund der Expansionsgeschwindigkeit seiner Schockwelle in einer bestimmten Region wurde RCW 86 nämlich auf ein Alter von etwa 10.000 Jahren geschätzt. Dieses Ergebnis wurde nun aufgrund von neuen Untersuchungen durch die beiden Röntgensatelliten Chandra und XMM-Newton in Frage gestellt. Anhand von genaueren Messungen der Synchrotronstrahlung in der nordöstlichen Region von RCW 86 gelangten Vink et al. (2006) zu einer direkten Abschätzung des Magnetfeldes in dieser Region, aus welchem sich dann die Expansionsgeschwindigkeit der Schockwelle gewinnen lässt. Diese wurde nun viel höher gemessen als in früheren Untersuchungen, womit auch das Alter der Supernova-Überreste automatisch geringer wird. Als eine obere Altersgrenze wurde etwa 2200 Jahre angegeben.
Wie kam es zu den unterschiedlichen Geschwindigkeitsabschätzungen? Vink et al. schlagen folgendes Szenario vor: Durch die stellaren Winde des ursprünglichen Sterns wurde vermutlich ein Teil des umliegenden Gases in eine gewisse Richtung gedrückt, womit sich um den Stern eine Art „Blase“ von verdichtetem Gas bildete. Trifft dann die Schockwelle auf diese Blase, wird sie relativ schnell gebremst. Hat diese Blase nun eine irreguläre Form, erhält die Schockfront je nach Ort eine andere Geschwindigkeit. Diese Überlegungen stehen in Übereinstimmung mit dem gemessenen Röntgenspektrum von RCW 86.
Durch sein geringes Alter wird eine Identifizierung von RCW 86 mit AD 185 wieder denkbar. Vink drückte sich in einem Interview vorsichtig aus (Bryner 2006): „Ich denke, es ist sehr interessant, dass wir nun mit einiger Zuversicht, jedoch nicht mit absoluter Sicherheit, sagen können, dass RCW 86 die Überreste von AD 185 sind.“
Bemerkenswert an dieser Geschichte ist, dass es offenbar ohne weiteres möglich ist, das Alter der Überreste einer Supernova von etwa 10.000 auf 2.000 Jahre umzudatieren. Das ist immerhin eine Korrektur von 80%. Damit wird beispielhaft deutlich, wie vorläufig wissenschaftliche Erkenntnisse in der Astrophysik sein können.
[Bryner J (2006) Astronomers Find Supernova First Spotted 2,000 Years Ago. www.space.com/scienceastronomy/060926_st_ancient_supernova.html; Rosado M, Le Coarer E & Marcelin M (1996) Kinematics of the galactic supernova remnants RCW 86, MSH 15-56 and MSH 11-61A. Astronomy & Astrophysics 315, 243-252; Vink J, Bleeker J, van der Heyden K, Bykow A, Bamba A & Yamazaki R (2006) The X-ray synchrotron emission of RCW 86 and the implications for its age. Astrophys. J. 648, 33-37.]
|

|
Weil sie so schön weithin sichtbar leuchten werden Supernovae (SN) gerne für Entfernungsmessung im Kosmos verwendet. Mit ihrer Hilfe bestimmt man Entfernungen von Galaxien im Bereich zwischen 10 Millionen und 10 Milliarden Lichtjahren.
Die ermittelten Entfernungen wiederum spielen beim Aufstellen kosmologischer Modelle eine Rolle. Allerdings eignen sich nicht alle Supernovae, sondern nur eine bestimmte Kategorie: die Supernovae Ia gelten als sogenannte kosmologische „Standardkerzen“, da bei einer solchen Supernovaexplosion im Wesentlichen immer derselbe Vorgang abzulaufen scheint. Ihre Verwendung zur Entfernungsmessung wird als sehr zuverlässig erachtet. Warum und wie das funktioniert, soll im Folgenden zunächst erläutert werden:
Gemäß dem gängigsten Modell zur Entstehung von Supernovae Ia zieht in einem Doppelsternsystem ein Weißer Zwerg Materie von seinem Begleiter ab, wodurch sich seine eigene Masse erhöht. Dieser Anstieg kann aber die sog. Chandrasekhar-Masse von ca.1,4 Sonnenmassen nicht übersteigen, denn sonst wird der Weiße Zwerg instabil. Durch den erhöhten Druck starten explosive thermonukleare Reaktionen, die den Stern schließlich ohne verbleibenden Rest völlig auseinander sprengen. Diese Explosion ist so hell, das sie eine ganze Galaxie überstrahlen kann, wobei die maximale Helligkeit der SN Ia erst nach gut zwei Wochen erreicht wird.
Dabei ist die Leuchtkraft einer SN Ia während ihres Intensitätsmaximums physikalisch vorgegeben. Bei der thermonuklearen Explosion entsteht u.a. das Isotop Nickel-56.
Der anschließende Zerfallsprozess dieses Isotops über Cobalt-56 zum stabilen Eisen-56 hält die hinausgeschleuderte Materie heiß und führt schließlich zum Intensitätsmaximum (Branch 2006). Die zeitlich unterschiedliche Zusammensetzung sichtbarer Elemente kann über eine sogenannte Lichtkurve verfolgt werden, d.h. das Lichtspektrum wird im Abstand weniger Tage über einen Zeitraum von mehreren Wochen bis Monaten aufgenommen. Anhand dieser Lichtkurve lassen sich Supernovae Ia von anderen Sternexplosionen unterscheiden. Berücksichtigt man noch die mögliche Absorption des Lichts auf dem Weg zu uns aufgrund von interstellarem Staub, kann daher aus der beobachteten scheinbaren Helligkeit einer SN Ia während des Intensitätsmaximums im Vergleich zur Leuchtkraft einer standardisierten SN Ia auf die Entfernung geschlossen werden.
Dieser Zusammenhang ist allerdings mit Fluktuationen behaftet; die Masse des erzeugten Nickel-56 schwankt und damit die maximale Leuchtkraft. Es besteht jedoch zusätzlich eine empirisch festgestellte Relation zwischen der maximalen Leuchtkraft und dem beobachteten Abklingen der Helligkeit des blauen Lichtanteils über die ersten 15 Tage nach dem Helligkeitsmaximum (Mazzali et al. 2007). Bei leuchtstärkeren SN Ia klingt die Lichtintensität langsamer ab als bei schwächeren (Filippenko 1997). Man verwendet daher die „Breite“ der Lichtkurve zur Kalibrierung der maximalen Leuchtkraft einer SN Ia. Die Brauchbarkeit von SN Ia zur Entfernungsmessung wurde zudem anhand einiger SN überprüft, die in Galaxien auftraten, deren Entfernungen bereits anderweitig bestimmt werden konnten, mit sehr guter Übereinstimmung (siehe z.B. Pailer & Krabbe 2006 und Literaturangaben darin).
Zwar gab es vereinzelt auch Ausreißer besonders heller (z.B. SN 1991T) oder schwacher (z.B. 1991bg) SN Ia, diese folgten jedoch dem bekannten Zusammenhang zwischen maximaler Leuchtkraft und Lichtkurve (Filippenko 1997), so dass ihre Entfernungen dennoch korrekt bestimmt werden konnten. Kürzlich sorgte dann aber jedoch ein in Nature erschienener Artikel (Howell et al. 2006) über eine wesentlich zu helle Supernova (SNLS-03D3bb bzw. SN 2003fg) für Irritationen. Zum einen lag ihr Helligkeitsmaximum beim 2,2-fachen einer durchschnittlichen SN. Zum anderen folgte sie nicht der üblichen Relation zwischen maximaler Leuchtkraft und Lichtkurve. Aus der Leuchtkraft kann man die Masse des Vorgängersterns abschätzen. Für SN 2003fg ergibt sich dabei eine zweifache (!) Sonnenmasse. Daraus ergeben sich zwei Konsequenzen.
Zum einen ist die Erklärung dieser speziellen Supernova eine Herausforderung für die theoretischen Modelle, da man bisher davon ausging, dass Weiße Zwerge nur bis zur 1,4-fachen Sonnenmasse stabil seien. SN Ia mit einem Vorgängerstern mit Super-Chandrasekhar-Masse kann man sich am ehesten in „jungen“ stellaren Umgebungen vorstellen. Das passt wiederum zum beobachteten Trend, wonach übermäßig helle SN Ia bevorzugt in solchen „jungen“ Umgebungen auftreten.
Zum anderen eignet sich SN 2003fg nicht als Standardkerze, solange die Ursachen für die Abweichung nicht verstanden und ggf. in ein revidiertes Modell für SN Ia integriert werden können. Durch ihre krasse Abweichung fällt sie momentan aber auf und wird einfach aussortiert. Möglicherweise existieren jedoch SN, deren abweichendes Verhalten nicht auffällig genug ist. Was passiert, wenn man sie übersieht? Man setzt möglicherweise ihre absolute Helligkeit über die Relation Leuchtkraft-Lichtkurve als zu niedrig an, was letztlich zu einer zu kleinen Entfernung führen würde (Branch 2006). Die Helligkeit nimmt über die Entfernung quadratisch ab. Würde man also zum Beispiel aus dem Helligkeitsverlauf von SN 2003fg auf ihre maximale Helligkeit schließen, erhielte man eine um den Faktor 1,45 (Wurzel aus 2,2) zu geringe Entfernung, d.h. ihre tatsächliche Entfernung ist knapp eineinhalb mal so groß wie die angenommene. Weniger drastische Ausreißer führen zu weniger drastischen Abweichungen. Dennoch könnte daraus ein Effekt für Entfernungsmessungen und somit für kosmologische Modellbildungen resultieren.
SN 2003fg befindet sich in einer Galaxie mit vergleichsweise hoher Rotverschiebung von z=0,24. Generell sind übermäßig leuchtstarke SN bevorzugt in jungen stellaren Umgebungen zu erwarten (s.o.). Mit zunehmender Rotverschiebung nimmt der Mittelwert des stellaren Alters ab. Damit geht dann eine Veränderung der durchschnittlichen Eigenschaften von Supernovae des Typs Ia einher. Weichen diese auch noch wie SN 2003fg vom bekannten Zusammenhang zwischen Lichtkurve und Leuchtkraft ab, so ist eine Kalibrierung nicht mehr möglich (Howell et al. 2006). Das könnte insofern zu einem Trend führen, indem man dabei für SN Ia mit hoher Rotverschiebung eine zu kleine Entfernung annimmt.
[Branch D (2006) Champagne supernova. Nature 443, 283-284; Filippenko AV (1997) Optical Spectra of Supernovae. Annu. Rev. Astron. Astrophys. 35, 309-355; Howell DA et al. (2006) The type Ia supernova SNLS-03D3bb from a super-Chandrasekhar-mass white dwarf star. Nature 443, 308-311; Mazzali PA, Röpke FK, Benetti S & Hillebrandt W (2007) A Common Explosion Mechanism for Type Ia Supernovae. Science 315, 825-827; Pailer N & Krabbe A (2006) Der vermessene Kosmos. Holzgerlingen, S. 39-42.] |
|
Bisher vermutete man, dass Zellen mit Zellkern, sogenannte Eukaryoten, im Laufe einer Evolution erst entstanden sind, nachdem Cyanobakterien die Atmosphäre mit Sauerstoff angereichert hatten. Dieses Ereignis liegt – nach herkömmlicher Zeitrechnung – 2,3 Milliarden Jahre zurück. Nach der Anreicherung von Sauerstoff in der Atmosphäre soll es anschließend eine globale ca. 100 Millionen Jahre lange extreme Eiszeit gegeben haben (Schneeball-Erde). Erst danach, vor etwa 1,7 Milliarden Jahren, sollen Eukaryoten auf der Weltbühne erschienen sein. Neue Beweise für das Vorhandensein von Eukaryoten vor diesen Zeitpunkten fanden sich in winzigen, in Quarz eingeschlossenen Öltropfen. Diese Öltropfen wurden analysiert und Buick und Mitarbeiter von der Washington Universität in Seattle fanden in Proben, die 2,4 Milliarden Jahre alt sind, Sterole, u.a. auch Cholesterol. Diese Moleküle sind biogenen Ursprungs und werden nahezu ausschließlich von Eukaryoten hergestellt. Diese Befunde haben im Rahmen der Evolutionstheorie mehrere Auswirkungen: Zum einen muss Sauerstoff auf der Erde schon lange vor der Oxygenierung der Atmosphäre produziert worden sein. Des Weiteren kann die Erde nicht so extrem abgekühlt sein, wie einige Modelle dies vorschlagen, und Eukaryoten betreten demnach die Bühne des Lebens zusammen mit den Archaea, nicht danach. Interessant ist auch noch die folgende Aussage von Buick: „Bisher dachte man, dass komplexe Kohlenwasserstoffe im Laufe der Zeit abgebaut werden. Die Tatsache, dass sie viele Milliarden Jahre überlebten, zeigt, dass wir auch auf dem Mars nach Leben suchen könnten.“ Was aber, wenn die Erde gar nicht so alt ist und die Biomoleküle nur deshalb intakt gefunden werden? Oder andersherum: Könnte man aus intakten Biomolekülen auf eine junge Erde schließen? Ähnliche Überlegungen gelten auch für DNA und gar lebende Bakterien aus Bernstein.
[Holzman D (2006) Eukaryotes apparently appeared far earlier, about when archaea arose. Microbe 1, 355-356.]
|
 |
| Abb.1: Zeichnerische und graphische Darstellung von Parapandorina aus der Doushantuo-Region Südchinas, nach Fotos aus Donoghue (2006), rechts nach einem Foto mit Röntgen-Mikrotomographie, mit zahlreichen Vakuolen. Eine neue Studie deckte einige Ähnlichkeiten mit Riesenbakterien auf. |
|
Die Deutung von Fossilien kann mitunter ausgesprochen schwierig sein, insbesondere wenn sie fast mikroskopisch klein sind. So wird durch eine neue Studie die Deutung bedeutsamer früher Tierfossilien aus dem oberen Präkambrium in Frage gestellt: Nach ihrer Entdeckung zunächst als älteste fossile Nachweise von Embryonen von Vielzellern interpretiert (Xiao et al. 1998), entpuppen sich Fossilien der Gattungen Parapandorina (Abb. 1) und Megasphaera aus der Doushantuo-Formation aus China nun vermutlich als Riesenbakterien (Bailey et al. 2006). Dies ergab sich durch einen Größen- und Proportions-Vergleich mit Thiomargarita, dem größten heute lebenden Bakterium, das in Sedimenten vor der Küste Namibias und im Golf von Mexiko entdeckt wurde. Thiomargarita ist etwa so groß wie die Doushantuo-Fossilien, und ist mit einer großen zentralen Vakuole und vielen kleinen Vakuolen ausgefüllt. Bei der Teilung entsteht ein Gebilde, das einem Vierzellstadium eines Vielzellers gleicht. Solche Teilungsstadien täuschen also im fossilen Erscheinungsbild wenigzellige Embryonen vor. Die Tatsache, dass solche Teilungsstadien in großer Zahl in den Doushantua-Phosphoriten vorkommen, jedoch keine ausgewachsenen Exemplare gefunden wurden, sperrt sich gegen die Deutung der Fossilien als Tierembryonen (Donoghue 2006, 155).
Dennoch kann der Fall nicht ad acta gelegt werden. Denn auch die Deutung von Parapandorina und Megasphaera als Riesenbakterien ist nicht in jeder Hinsicht stimmig: Beispielsweise sind von Thiomargarita nur maximal 3 Abfolgen einer Zellteilung (8 zusammenhängende Zellen) bekannt, während bei Parapandorina Aggegrate von über 100 Zellen bekannt sind, was mindestens 7 Teilungsstadien erfordert. Die Klassifikation der Fossilien kann daher nicht als geklärt gelten (Donoghue 2006, 156). Es bleibt dabei, was Donoghue (2006) einleitend in seinem Kommentar feststellte: „Der Ursprung der Tiere ist fast ein solches Geheimnis wie der Ursprung des Lebens.“ Wie weit die evolutionäre Geschichte des tierischen Lebens ins Präkambrium reiche, sei reich an Spekulationen und spärlich an Belegen.
[Bailey JV, Joye SB, Kalanetra KM, Flood BE & Corsetti FA (2006) Evidence of giant sulphur bacteria in Neoproterozoic phosphorites. Nature 445, 198-201; Donoghue PCJ (2006) Embryonic identity in crisis. Nature 445, 155-156; Xiao S, Zhang Y & Knoll AH (1998) Three-dimensional preservation of algae and animal embryos in a Neoproterozoic phosphorite. Nature 391, 553-558.]
|
|
Gliederfüßer mit sechs Beinen (Hexapoda), darunter als Hauptgruppe die Insekten, sind die vielseitigste Gruppe unter den heutigen Lebewesen, sowohl was ihre Artenzahl als auch was ihre Lebensräume betrifft. Dennoch wird über ihren Ursprung kontrovers diskutiert. Mit diesen Feststellungen beginnen Glenner et al. (2006) einen Kurzbeitrag über den Ursprung der Insekten in Science. Als „Schlüsselproblem“ betrachten sie die fast vollständige Abwesenheit von Fossilien, welche die Hexapoden mit den anderen größeren Gliederfüßergruppen verbinden. Dazu gehören die Crustaceen (Krebsartige), Myriapoden (Hundert- und Tausendfüßer) und die Cheliceraten wie Skorpione und Spinnen. Während die Crustaceen schon im Oberkambrium (vor ca. 510 Millionen Jahren nach üblicher Datierung) fossil überliefert sind, tauchen die Hexapoden erst 100 Millionen Jahre später ab dem Devon auf.
Auch die Merkmalsverteilungen dieser Gruppen legen offenbar keine eindeutigen Verwandtschaftsbeziehungen nahe: die Hexapoden wurden im Laufe der Zeit mit allen eben genannten Gliederfüßergruppen schon stammesgeschichtlich verbunden – ein sicherer Hinweis auf die mosaikartige bzw. baukastenartige Verteilung von Merkmalen, die sich gegen ein klares Abzweigungsschema sperrt.
Traditionell wurden die Hexapoden mit den Myriapoden aufgrund morphologischer (gestaltlicher) Ähnlichkeiten zusammengefasst. Molekulare Daten und neuere morphologische Studien dagegen deuten auf eine Verwandtschaft der Hexapoden mit den Crustaceen hin, während neurologische Studien eine Verbindung der Myriapoden mit den Cheliceraten nahelegen. Insgesamt zeigt sich einmal mehr, dass morphologische und molekulare Daten nicht ohne weiteres zusammenpassen und sich nicht gegenseitig stützen.
Die durch die neueren Studien gewonnenen Daten ermöglichen nun aber ein neues evolutionäres Szenario der Entstehung der Hexapoden. Sie könnten von einer vornehmlich im Süßwasser lebenden Untergruppe der Crustaceen, den Branchioden (zu denen z. B. Wasserflöhe gehören) abgeleitet werden. Diese stammesgeschichtliche Beziehung würde auch gut zum zeitlichen Auftreten in der Fossilüberlieferung passen. Allerdings gibt es auch hier einen Widerspruch zwischen morphologischen und molekularen Daten: Erstere verbinden die Hexapoden eher mit den Krabben und Flusskrebsen, letztere eher mit den genannten Branchiostomen, die aber bezüglich der Fossilberlieferung besser passen. Die Hexapoden sollen demnach relativ spät im Süßwasser entstanden sein und von dort aus das Land erobert haben,während sie nach bisherigen Vorstellungen schon viel früher unter marinen (meerischen) Verhältnissen im Kambrium entstanden sein sollen.
Dass die auf Land so unvergleichlich erfolgreichen Hexapoden nicht mehr die Wasserlebensräume erobern konnten, wird darauf zurückgeführt, dass dieser Lebensraum schon besetzt gewesen sei. Evolutionstheretisch gedacht erscheint dies aber angesichts des ungeheuren „Erfolgs“ der Hexapoden mindestens zweifelhaft und eine ad-hoc-Annahme. Das neue Entstehungsszenario ändert auch nichts an den markanten fossilen Lücken wie auch den eingangs erwähnten Merkmalswidersprüchen.
[Glenner H, Thomson PF, Hebsgaard MB, Sorensen MV & Willerslev E (2006) The Origin of Insects. Science 314, 1883-1884.]
|
|
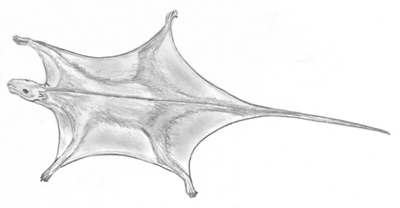
Die Fähigkeit zum Gleitflug findet sich in einer ganzen Reihe von Tiergruppen, so bei Reptilien, Fröschen, Fischen und Säugern. Zur Geschichte des Gleitflugs bei jenen Säugetieren lieferten Meng et al. kürzlich interessante neue Ergebnisse. Mindestens siebenmal unabhängig soll die Gleitfähigkeit allein bei Säugetieren – auf vier verschiedene Säugetierordnungen verteilt – entstanden sein – ein bemerkenswertes Beispiel von Konvergenz. Dabei galt bislang die Gruppe der Fledertiere als die erste, die den Luftraum erobert haben soll, und zwar im Untereozän (ca. 50 Millionen Jahre nach radiometrischen Datierungen). Nagetiere mit der Fähigkeit zum Gleitflug tauchen fossil erst vor ca. 30 Millionen Jahren auf. Es schien daher eine klare Sache zu sein, dass die Eroberung des Luftraumes durch Säuger erst recht spät, in der Erdneuzeit, vonstatten gegangen war.
Ein Team US-amerikanischer und chinesischer Wissenschaftler berichtete nun jedoch in Nature über einen spektakulären Fund, der ein erhebliches Umschreiben der Geschichte der Flugfähigkeit unter Säugern erfordert: ein zum Gleiten befähigtes Säugetier aus der Unterkreide. Volaticotherium antiquum (etwa „altes Flugtier“) heißt das gerade einmal 12-14 cm lange und 70g schwere, heutigen Flughörnchen ähnelnde und sich allem Anschein nach von Insekten ernährende Tier, dessen Fossil bei Daohugou in der Inneren Mongolei Chinas gefunden wurde (vgl. Abb. 1). (Die Namensbezeichnung wurde nachträglich von antiquus auf antiquum geändert: Nature 446, 102.) Die von den Forschern zwischen Vorder- und Hinterextremitäten ausgemachte, dicht mit Haaren besetzte Struktur wurde als Flughaut identifiziert. Mit einem geologischen Alter von 125 Millionen Jahren beweist dieser Fund, dass schon im Erdmittelalter durch die Luft gleitende Säugetiere lebten. Völlig unabhängig von der Entstehung der Fledermäuse und Flughunde hätten damit einige Kleinsäuger folglich schon mindestens 70 Millionen Jahre früher die Fähigkeit des Gleitfluges erworben.
Der Fund demonstriert einmal mehr, dass frühe Säuger in ihren Fortbewegungsstrategien und Lebensstilen äußerst verschiedenartig waren. Diese erstaunliche Diversifikation in einem solch frühen Stadium der Geschichte der Säugetiere war evolutionstheoretisch früher nicht erwartet worden und reiht sich ein in eine ständig länger werdende Liste von Bauplantypen, die bereits zu Zeiten der „primitiven“ Säuger hochspezialisiert waren (Martin 2006; Luo & Wible 2005; vgl. Junker 2005; Junker & Scherer 2006, 230f.).
Bemerkenswert ist nicht nur, dass durch diese Entdeckung eine neue Ordnung („Volaticotheria“) geschaffen wurde; sie wirft auch eine in der Schulwissenschaft bislang unangebrachte Frage auf: Waren es tatsächlich die Vögel oder nicht doch sogar die Säuger, welche den Luftraum als erste eroberten? Das häufige unabhängige (konvergente) – und nun auch äußerst frühe – Auftreten der Fähigkeit zum Gleitflug stellt evolutionstheoretische Deutungen auf eine harte Probe.
[Martin T (2006) Early Mammalian Evolutionary Experiments. Science 311, 1109-1110; Luo ZX & Wible JR (2005) A late Jurassic digging mammal and early mammalian disversification. Science 308, 103-107; Meng J et al. (2006) A Mesozoic gliding mammal from northeastern China. Nature 444, 889-893; Junker R & Scherer S (2001) Evolution – ein kritisches Lehrbuch; Junker R (2005) Alte Termitenfresser. Stud. Int. J. 12, 88.]
|
|
Neunaugen (Petromyzontida) sind fischähnliche Wirbeltiere mit aalartigem langgestrecktem Körper. Zusammen mit den Schleimfischen (Myxinoidea) werden sie zu den Rundmäulern (Cyclostomata) zusammengefasst, die insgesamt ca. 60 Arten umfassen. Sie besaßen keine Kiefer und auch keine paarigen Flossen, sondern einen flossenartigen Rücken- und Schwanzsaum. Unter anderem aus diesen Gründen gelten sie als stammesgeschichtlich primitiv. Andererseits sind sie ausgesprochen spezialisiert und leben als Parasiten, die sich mit ihrem rundlichen, mit Hornzähnen gesäumten Maul an Fischen festaugen, Blut trinken und Fleischstücke herausraspeln. Der Name „Neunauge“ entstand durch den augenartigen Eindruck, den die Nasenöffnung und die sieben seitlichen Kiemenspalten erwecken. Fossilien von ihnen sind kaum bekannt, die bislang ältesten Formen beider Untergruppen stammten aus dem Karbon.
Bis zum Ende des 20. Jahrhunderts wurden die Rundmäuler als „degenerierte“ Abkömmlinge der gepanzerten Kieferlosen (als Ostracodermi zusammengefasst) betrachtet, die aus dem Ordovizium bis zum Devon fossil überliefert sind (Janvier 2006, 921).
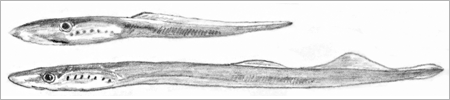 |
| Abb.1: Reconstruktion of Priscomyzon riniensis (oben) im Vergleich mit dem heute lebenden Neunauge Lampetra fluviatilis (unten). (Nach Gess et al. 2006) |
|
Nun haben Wissenschaftler ein gut erhaltenes Fossil eines Neunauges (Priscomyzon riniensis) in marinen (meerischen) Ablagerungen des Oberdevons in Grahamstown/Südafrika gefunden (Gess et al. 2006; Abb. 1). Es weist alle typischen spezialisierten Merkmale heutiger Neunaugen auf und unterscheidet sich von diesen nur in geringfügigen Details. Der Körperbau der Neunaugen erweist sich damit als außerordentlich stabil, was für einen Zeitraum von 360 Millionen Jahren im evolutionstheoretischen Deutungsrahmen „erstaunlich“ ist (Janvier 2006, 923) – es handelt sich um ein ausgeprägtes „lebendes Fossil“. Hardistiella montana aus dem Unterkarbon der USA war zuvor der geologisch früheste Angehörige der Neunaugen (Thenius 2000, 118). Damit wird das Auftauchen des evolutionstheoretisch ältesten Wirbeltiers, das mit seinen heutigen Verwandten praktisch identisch ist, geologisch noch einmal zurückverlegt.
Mit diesem unerwartet frühen fossilen Auftreten müssen nun die Vorstellungen über die Abstammungsverhältnisse deutlich revidiert werden. Denn der Ursprung der Neunaugen muss soweit vorverlegt werden, dass die bisherige Ableitung von den kieferlosen Ostracodermen aufgegeben werden muss. Ihr Ursprung bleibt somit im Dunkeln.
Janvier weist in seinem Kommentar darauf hin, dass die Verwandtschaftsbeziehungen zwischen den heutigen Schleimfischen, Neunaugen und kiefertragenden Wirbeltieren „heiß debattiert“ wird, weil gestaltliche und physiologische Merkmale mit Sequenzdaten von DNA und RNA im Konflikt stehen – einmal mehr, möchte man ergänzen. Die anatomischen Merkmale sprechen für eine nähere Verwandtschaft der Neunaugen mit den kiefertragenden Wirbeltieren, während die molekularen Daten die Neunaugen und die Schleimfische als Schwestergruppen nahelegen. Das nun entdeckte Fossil kann hier nicht weiterhelfen, da es keine neuen Merkmalskombinationen aufweist.
Der praktisch identische Körperbau der Neunaugen seit dem Devon ist in der Tat erstaunlich (so Janiver; s.o.), denn während ihrer nachgewiesenen Lebenszeit hat sich im Sinn von Evolution mehrfach eine neue Fischwelt entwickelt und ist ganz überwiegend wieder verschwunden. Um nur die bedeutendsten, sehr vielgestaltigen Großgruppen zu nennen: Neben den (schon erwähnten) paläozoischen panzertragenden, kieferlosen Schalenhäutern (Ostracodermi) mehrere paläozoische Fischgruppen wie Stachelhaie (Acanthodii) oder gepanzerte Plattenhäuter (Placodermi). Sodann die jungpaläozoisch-mesozoischen Knorpelganoiden (Chondrostei) und mesozoischen Knochenganoiden (Holostei), die wiederum durch die in Trias und Jura auftauchenden „modernen“ Knochenfische (Teleostei) – endgültig in der Kreide – abgelöst wurden. Auch Quastenflosser (Crossopterygii) und Lungenfische (Dipnoi), etwas später auch Haie (Selachii) oder Seedrachen (Chimären) treten mit Formen auf, die ihren heutigen Nachkommen sehr ähneln, aber bei keiner Fischgruppe ist dieses Phänomen so auffällig wie bei Neunaugen (und Schleimfischen). An ihnen ging der Selektionsdruck, der in den gleichen Biotopen die genannten, sehr verschiedenen Fischwelten hervorgebracht haben soll, am längsten vollständig vorbei – kein gutes Indiz für ein umfassendes Vermögen der Selektion. Wenn also die Selektion die Neunaugen nicht nennenswert verändert hat, war dann Selektion überhaupt die Triebfeder, die zur gleichen Zeit zu den verschiedenen Fischgruppen geführt hat?
[Gess RW, Coates MI & Rubidge BS (2006) A lamprey from the Devonian period of South Africa. Nature 443, 981-984; Janvier P (2006) Modern look for ancient lamprey. Nature 443, 921-924; Thenius E (2000) Lebende Fossilien. München]
|
|
Wenn eine Art in neue Lebensräume verschleppt wird oder sie erobert, muss sie in der Regel neue Lebensbedingungen bewältigen. Oder die bisher bereits einheimischen Arten stehen beim Eindringen neuer Arten vor entsprechenden Herausforderungen. Und die Reaktion bzw. Anpassung muss schnell erfolgen, sonst ist das Überleben ernsthaft gefährdet. Dies ist nur möglich, wenn die Arten eine gewisse Flexibilität mitbringen. Ein schönes Beispiel dafür liefern Miesmuscheln in Neuengland (Nordosten der USA), die mit Krabben fertig werden mussten, die aus Asien eingeschleppt und seit 1988 an den Küsten von New Jersey beobachtet worden waren. Von dort aus breiteten sie sich nach Norden aus. Die Muscheln waren in der Lage, in kurzer Zeit ihre Schalen dicker werden zu lassen. Auf eine unbekannte Weise, vielleicht durch Wahrnehmung chemischer Substanzen, merken die Muscheln, wenn die Krabben in ihre Nähe kommen, und verdicken daraufhin im Laufe einiger Monate ihre Schalen um 5-10% (Freeman & Byers 2006; Stokstad 2006). Das genügt, um den Krabben das Öffnen der Schalen soweit zu erschweren, dass sie den Öffnungsversuch häufiger aufgeben, um sich leichterer Beute zuzuwenden. Dies war schon früher bei einer bereits um 1800 eingeschleppten Krabbenart festgestellt worden. Nun zeigte sich, dass die Muscheln binnen 15 Jahren „lernten“, sich durch Verdickung ihrer Schalen des neuen Räubers zu erwehren.
Die Untersuchungen von Freeman & Byers geben den Biologen Hoffnung, dass das Einschleppen von Arten nicht notwendigerweise eine massive Gefährdung der einheimischen Lebewelt bedeuten muss; die einheimischen Arten sind offenbar nicht unbedingt hilflos. Solche Befunde passen gut zum Konzept polyvalenter Stammformen, einem Konzept, das im Rahmen der Schöpfungslehre vertreten wird. Demnach weisen die Stammformen der heutigen Grundtypen ein vielseitiges Erbgut auf, das ihnen Anpassungsfähigkeit und Flexibilität verleiht.
[Freeman AS & Byers JE (2006) Divergent Induced Responses to an Invasive Predator in Marine Mussel Populations. Science 313, 831-833; Stokstad E (2006) Native mussel quickly evolves fear of invasive crab. Science 445, 745.]
|
|
Der Grundtyp der Hundeartigen bietet ein breites Spektrum an Variationen und eine besonders augenfällige Vielfalt auf phänotypischer (gestaltlicher) Ebene. Dass die zum Teil sehr großen morphologischen Unterschiede (Unterschiede in der äußeren Erscheinung) jedoch nicht auf ebenso ausgeprägte Unterschiede im Erbgut zurückzuführen sind, demonstriert eine neue Studie von Sutter (2006), wonach einem einzigen Gen, Igf-1 (insulin-like growth factor 1), eine entscheidende Bedeutung für die Körpergröße der Hunde zukommt. Zu diesem Ergebnis kamen Sutter und sein Team, nachdem sie 500 portugiesische Wasserhunde aufgrund von Röntgenaufnahmen und Skelettmessungen in die Kategorien „groß“ und „klein“ aufgeteilt und nach zwischen den beiden Gruppen bestehenden Erbgutunterschieden gesucht hatten (Check 2006).
 |
| Abb.1: Groß oder klein – nur ein kleiner genetischer Unterschied? |
|
Die Sequenzierung und Veröffentlichung des Hundegenoms waren bereits im Vorjahr von einem Team, dem auch Sutter angehört hatte, vorgenommen worden. Diese Datenbasis erleichterte die Suche der Forscher sehr, weil das Igf-1-Gen im Gefolge der Sequenzierung bereits als eine der wenigen Gensequenzen erkannt worden war, die innerhalb einer Art stark variieren. Außerdem war bereits bekannt, dass dieses Gen bei Mäusen einen Einfluss auf die Körpergröße hat, da diese bei Ausfall dieses Gens sehr kleinwüchsig bleiben. Um zu testen, ob derselbe Zusammenhang auch bei den Hunden besteht, wurden 350 Individuen unterschiedlicher Rassen und sehr unterschiedlicher Größe im Hinblick auf dieses Gen miteinander verglichen. Dabei zeigte sich, dass nahezu alle der 18 kleinen Hunderassen, die für das Experiment herangezogen worden waren, genau dieselbe Igf-1-Gen-Variante besaßen, während diese Gen-Variante bei den 15 großen Rassen praktisch gar nicht vorkam.
Bei der Züchtung der kleinen Hunderassen kreuzte der Mensch die jeweils kleinen Individuen, welche die für die geringe Körpergröße verantwortliche Variante des Gens besaßen. Dadurch wurde diese Genvariante im Laufe dieses Selektionsprozesses in den kleinen Hunderassen fixiert.
Die Ergebnisse zeigen, dass bereits geringe genotypische Veränderungen für phänotypische Vielfalt innerhalb eines Grundtyps sorgen können.
Interessant ist auch, dass angenommen wird, diese Gen-Variation sei bereits sehr früh in der Geschichte der Diversifikation der Hundeartigen evolviert und habe somit schon früh zur Selektion und somit zur Spezialisierung der Hunderassen bereitgestanden. Dies passt sehr gut zur Annahme polyvalenter Stammformen, welche schon zu Beginn des Aufspaltungsprozesses ein breites Variationsspektrum besaßen (vgl. Junker & Scherer 2006, 297ff.).
[Check E (2006) One gene between tiny dogs and giant ones? http://www.nature.com/news/2006/061009/full/061009-12.html; Junker R & Scherer S (2006) Evolution – ein kritisches Lehrbuch. Gießen, 6. Auflage]
|
|
Spätestens seit Moby Dick weiß jeder, dass Wale gar nicht so dumm sind, wie man von diesen recht schwerfällig anmutenden Tieren vielleicht gedacht hätte. Mittlerweile konnte durch wissenschaftliche Studien nachgewiesen werden, dass diese Tiere extrem intelligent sind. So sagt z.B. Patrick R. Hof (2006) von der Mount Sinai School of Medicine in New York: „Sie kommunizieren durch umfangreiche Repertoires an Gesängen, erkennen ihre eigenen Gesänge und erfinden neue. Außerdem schließen sie sich zu Gruppen zusammen, um Jagdstrategien zu planen, bringen diese den jüngeren Tieren bei und haben soziale Netzwerke entwickelt, die denen von Menschen und Affen ähneln …“
Kürzlich haben Forscher herausgefunden, dass die Wale nicht nur ähnliche Verhaltensweisen wie Menschen und Affen aufweisen, sondern dass dies auch auf neurologischer Ebene begründet ist. So berichtete Michael Balter in Science von der Entdeckung spezieller Nervenzellen (sog. Spindelneuronen) in den Gehirnen mancher Wale. Stark vereinfacht kann man sagen, dass diese Nervenzellen in Hirnregionen, in denen Emotionen verarbeitet werden, eine rasche intuitive Beurteilung von Situationen gewährleisten.
So weit, so gut. Wirklich interessant wird das ganze erst, wenn man die Vorgeschichte kennt: Diese begann mit einem Artikel von Helen Phillips in New Scientist aus dem Jahr 2004, in dem „The cell that makes us human“ (Die Zelle, die uns menschlich macht) beschrieben wurde. Unter diesem Titel wurde außerdem behauptet: „Die Entdeckung eines Nerventyps, der nur bei Menschen und unseren nächsten Verwandten vorkommt, erregt Aufsehen.“ Es passte alles wunderbar – eine gemeinsame Nervenzelle, ein gemeinsamer Vorfahr. Diese Nervenzellen, entdeckt vom Hirnforscher John M. Allman vom California Institute of Technology in Pasadena und seinem Team, sind ziemlich groß und zigarettenförmig aufgebaut und vernetzen verschiedenste Hirnregionen miteinander. Menschen besitzen laut Allmans Analyse gut 80 000 solcher Spindelneuronen, Gorillas 16 000 und Schimpansen 1800.
Allman et al. (1999) hatten festgestellt, dass die Spindelzellen in keiner anderen Primatenart und keiner anderen Säugergruppe entdeckt worden seien und dass ihr Volumen mit einem Maß der Enzephalisation höherer Primaten korreliere. Sie schreiben: „Diese Beobachtungen sind von besonderem Interesse für die Betrachtung der Evolution des Neocortex der Primaten, da sie mögliche adaptive Veränderungen und funktionelle Modifikationen während der letzten 15-20 Millionen Jahre im vorderen cingulären Cortex aufzeigen, einer Region, die eine größere Rolle in der Regulation vieler Aspekte der autonomen Funktion und bestimmter kognitive Vorgänge spielt.“
Die neuen neurologischen Erkenntnisse über die Spindelzellen bei Walen bestätigen die oben kurz geschilderten Ergebnisse der Verhaltensforschung und haben darüber hinaus die Diskussion über die Rechtfertigung der Zufügung von Leid beim Walfang neu entfacht.
Darüber hinaus wirft die Entdeckung der Spindelzellen bei Walen jedoch noch eine ganz andere Frage auf – die Frage, wie das innerhalb des Evolutionsparadigmas zu verstehen sei. Erst kürzlich berichtete Junker (2006) von einer „bitteren Konvergenz“: Teilen Affen und Menschen ein Merkmal, wird dies praktisch automatisch als Homologie, als gemeinsames Erbe eines gemeinsamen Vorfahrens interpretiert. In dem von Junker genannten Fall musste diese Interpretation jedoch aufgegeben werden: die Sensibilität für den Bitterstoff Phenylthiocarbamid (PTC) soll nun vielmehr zweimal unabhängig, bei Menschen und Affen, entstanden sein – eine im wahrsten Sinne des Wortes „bittere“ Konvergenz.
Mit den neuen Ergebnissen bezüglich der Spindelzellen scheint sich nun eine „nervige“ Konvergenz in die lange Liste der erstaunlichen Konvergenzen im Organismenreich einzureihen (vgl. den Beitrag über das konvergente Auftreten des Gleitflugs auf Seite 48 dieser Ausgabe).
Wohl oder übel müssen Vertreter der Evolutionslehre annehmen, Spindelneuronen hätten sich im Laufe der Evolution bei Walen und Menschen völlig unabhängig voneinander entwickelt, einmal vor etwa 30 Millionen Jahren und einmal vor 15 Millionen Jahren. Aufgrund der Verteilung des Besitzes von Spindelneuronen muss darüber hinaus für das Auftreten dieses Zelltyps bei den Walartigen entweder ein mehrfacher Verlust, ausgehend von einem gemeinsamen, Spindelzellen besitzenden Vorfahren angenommen werden, oder aber wiederum eine mehrfache Entstehung bei den zwei unterschiedlichen Unterordnungen der Walartigen. Außerdem ist es gut möglich, dass – evolutionstheoretisch gesehen – ein Teil dieses Prozesses ablief, als die Spindelzellen gerade auch das erste mal in der Vorfahrenlinie der Menschen auftauchten – was zusätzlich ein besonders seltener Fall einer parallelen Evolution darstellen würde.
Andererseits stützen diese Ergebnisse die Deutung des Auftretens von Konvergenzen als Design-Signal. Die freie Wiederverwendung gleicher bzw. sehr ähnlicher Strukturen bei sonst nicht näher verwandten Organismen passt gut in die Vorstellung eines uneingeschränkt agierenden Designers, der gleichsam ein „Baukastensystem“ frei nutzen kann (vgl. ReMine 1993; ReMine 2006; Junker 2000).
[Allman JM et al. (1999) A neuronal morphologic type unique to humans and great apes. Proc. Natl. Acad. Sci. 96, 5268-5273; Balter M (2006) Well-Wired Whales. Science NOW, http://sciencenow.sciencemag.org/cgi/content/full/2006/1127/1; Hof PR & van der Gucht E (2006) Structure of the cerebral cortex of the humpback whale, Megaptera novaeangliae (Cetacea, Mysticeti, Balaenopteridae). The Anatomical Record Part A: doi: 10.1002/ar. a. 20407; Junker R (2006) Bittere Konvergenz. Stud. Int. J. 13, 99; Junker R (2000) Ähnlichkeiten, Rudimente, Atavismen. Holzgerlingen; Phillips H (2004) The cell that makes us human. New Scientist 182, No. 2452, 32-35; ReMine WJ (1993) The Biotic Message.Evolution versus Message Theory. Saint Paul; ReMine WJ (2006) Evidence for message theory: a review of Life’s Solution: Inevitable Humans in a Lonely Universe by Simon Conway Morris. Journal of Creation 20(2), 29-35.]
|
|  |