 |
Konvergente Kooption von Pax-Genen
Oder: Was einmal taugt, taugt auch mehrmals
von Reinhard Junker
Studium Integrale Journal
18. Jahrgang / Heft 1 - April 2011
Seite 38 - 40
|
 |
Zusammenfassung: Eine der überraschenden Entdeckungen der letzten Jahre ist die vielfache Verwendung derselben Gene in verschiedenen Zusammenhängen. Evolutionstheoretisch wird postuliert, dass Gene oft „zweckentfremdet“ wurden; man spricht von „Kooption“ oder „Rekrutierung“. Immer häufiger zeigt sich darüber hinaus, dass ähnliche oder auch unähnliche Gene mehrfach unabhängig für den selben Zweck kooptiert wurden. Wie kann es sein, dass dies durch einen Evolutionsvorgang erfolgt, der nicht an irgendwelchen Zielen orientiert ist, aber dennoch zu einem sehr ähnlichen Ergebnis gelangt? Beispielsweise muss nach neuen Befunden die mehrfache Kooption von Pax-Genen angenommen werden. Diese Gene spielen eine herausragende Rolle bei der ontogenetischen (individuellen) Entwicklung der Augen. Eine neue Untersuchung darüber lässt die erheblichen Probleme aufscheinen, die sich einem natürlichen Evolutionsprozess entgegenstellen.
| |
|
Einführung
Bei der Ausbildung von Augen spielt eine Reihe von Steuergenen eine wichtige Rolle. Das bekannteste darunter ist Pax-6. Dieses Gen kontrolliert die Augenentwicklung auf höchster Ebene bei den Bilateria („Zweiseitentiere“, dazu gehören z. B. die Gliederfüßer und die Wirbeltiere). Pax-6 ist z. B. beim Linsenauge der Säugetiere nötig, um eine nachfolgende Kaskade von etwa 2500 Genen in Gang zu setzen, die für die Formbildung des Auges wichtig sind (Halder et al. 1995); es wird daher auch als Masterkontrollgen bezeichnet. Pax-6 ist bei Arten, die zu ganz verschiedenen Tierstämmen gehören, so ähnlich, dass man es ohne Probleme zwischen sehr verschiedenen Tieren austauschen kann. So konnte die Arbeitsgruppe von Walter Gehring bereits 1995 zeigen, dass das Pax-6-Gen der Maus (welches hier eyeless heißt) auch in Fliegen seine Wirkung entfalten kann und die Bildung eines (Fliegen-)Auges auslöst (Halder et al. 1995).
Aufgrund dieser Ähnlichkeit wurde zunächst für alle Tiere mit Augen ein Vorfahr mit einem Urauge postuliert (Gehring 1996), dessen ontogenetische Bildung bereits mittels Pax-6 gesteuert wurde (vgl. Abb. 1). Da das Komplexauge der Insekten und das Linsenauge der Wirbeltiere in ihrem Aufbau grundverschieden sind, muss das Urauge dieses postulierten gemeinsamen Vorfahren so einfach gedacht werden, dass es als Basis für die evolutionäre Entstehung beider Augentypen dienen kann. Bei einer völlig unabhängigen Entstehung von Komplexauge und Linsenauge müsste man alternativ annehmen, dass das Steuergen Pax-6 erst sekundär und unabhängig aus einem anderen genetischen Funktionszusammenhang für die Augenentwicklung verwendet wurde, nachdem Komplex- und Linsenaugen evolutionär bereits entstanden waren. Dieser Prozess wird als extrem unwahrscheinlich betrachtet.
|
|
|
Kooption
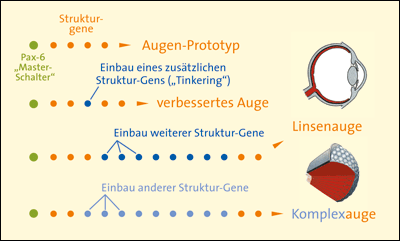 |
| Abb. 1: Stark schematisierte Darstellung der Entstehung verschiedener Augentypen, ausgehend von einem hypothetischen Urauge, in dessen ontogenetischer Entwicklung bereits das Masterkontrollgen Pax-6 beteiligt war. Die hypothetische evolutive Entwicklung erfolgt durch sukzessive Einfügung weiterer Gene, die ebenfalls unter der Kontrolle von Pax-6 stehen. Pax-6 soll ursprünglich aus einem anderen Kontext für die Augenentwicklung rekrutiert („zweckentfremdet“) worden sein. |
|
Der hypothetische Vorgang der Inanspruchnahme vorhandener Gene für eine neue Aufgabe wird als „Kooption“ bezeichnet; man spricht auch von Gen-Rekrutierung oder auch von einer „Neuverschaltung“ von Genen. Die damit verknüpften Prozesse bzw. die einer solchen Neuordnung zugrunde liegenden Mechanismen sind aber weitgehend hypothetisch.
Da Kooption evolutionstheoretisch sehr unwahrscheinlich ist, muss man annehmen, dass Pax-6 schon sehr früh in der mutmaßlichen Evolution der Augen als sogenanntes Masterkontrollgen in Erscheinung trat, zu einem Zeitpunkt, als das Urauge noch sehr einfach und unspezialisiert gebaut war (s. o.; vgl. Abb. 1).
Aufgrund der phylogenetischen Analysen (Ermittlung von Abstammungsverhältnissen durch Merkmalsvergleiche) stellte sich heraus, dass angesichts der genetischen Merkmalsverteilungen (hier Pax-Gene) zweifache oder sogar mehrfache konvergente Kooptionen gleicher Gene angenommen werden müssen. Das heißt: Die gleichen oder ähnlichen (Pax-)Gene wurden zweimal oder mehrfach unabhängig in unterschiedlichen stammesgeschichtlichen Linien (Hydrozoa, Cubozoa, Bilateria) in gleicher Weise für eine identische Funktion rekrutiert. Angesichts der Schwierigkeiten, die sich schon bei einer erfolgreichen Integration eines Gens in einen anderen Funktionszusammenhang ergeben, wirft die Erklärung einer mehrfach unabhängigen gleichartigen Neu-Inanspruchnahme desselben Gens erst recht Fragen nach den Mechanismen auf. Mit der Zunahme der zu postulierenden Kooptionen (wie sie z. B. auch bei den Crystallinen für die Augenlinsen notwendig ist) wurde mehr und mehr von der Vermutung einer monophyletischen Entstehung der Augen Abstand genommen (Gregory 2008, Piatigorsky 2008).
|
|
Fünf Pax-Unterfamilien
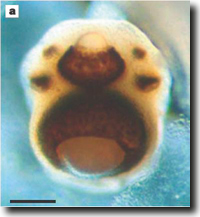 |
| Abb. 2: Die Augen der zu den Würfelquallen gehören Art Tripedalia cystophora: Obere und untere Linse, flankiert von zwei Paaren einfacherer Augen. Balken: 0,1 mm. (Aus Nilsson et al. 2005, Abdruck mit freundlicher Genehmigung) |
|
Die Vertreter der Pax-Gengruppe werden in fünf Unterfamilien unterteilt. Pax-Gene besitzen in vielen Organismen zentrale Steuer- bzw. Schalterfunktionen in der Ontogenese verschiedenster Organe. Wie oben erwähnt, ist Pax-6 für die Augenentwicklung bei den Bilateria (Zweiseitentiere) entscheidend. Pax-6 kommt aber bei den Nesseltieren (Cnidaria) nicht vor. Interessanterweise haben vier der fünf Klassen der Nesseltiere, zu denen auch drei Klassen von Quallen gehören, Augen, z. T. sogar hochentwickelte Linsenaugen (Abb. 2). Kürzlich veröffentlichten Suga et al. (2010) eine Untersuchung über das Vorkommen von Pax-Genen bei der Qualle Cladonema radiatum aus der Klasse der Hydrozoa. In diesem Tier wurden drei Pax-Gene, jeweils einer anderen Unterfamilie angehörig, Pax-A, Pax-B und Pax-E, gefunden. Suga et al. (2010) konnten zeigen, dass Pax-A bei der Bildung der Linsenaugen von Cladonema eine führende Rolle zukommt. Bei Tripedalia, die zu der Klasse Cubozoa (Würfelquallen) gehört, wurde in einer früheren Untersuchung Pax-B als verantwortlich für die Augenentwicklung nachgewiesen (vgl. Abb. 3). Bei den Bilateria, den Hydrozoa und den Cubozoa werden also Pax-Gene aus verschiedenen Pax-Unterfamilien verwendet.
Suga et al. vermuten, dass die verschiedenen Pax-Unterfamilien durch Genduplikationen in einer sehr frühen Phase der Evolution aus einem urtümlichen Pax-Genvorläufer in einem augentragenden gemeinsamen Vorfahren entstanden sind, spätestens vor der Trennung der Bilateria und der Cnidaria (Abb. 3). Es sei daher sehr wahrscheinlich, dass in den drei Linien der Hydrozoa, Cubozoa und Bilateria diese drei verschiedenen Pax-Gene zunächst unterschiedlich erhalten blieben (Pax-A und Pax-B fehlt bei den Bilateria, Pax-6 bei den Cnidariern) und dann unabhängig für die jetzige analoge Aufgabe in der strukturellen Augenentwicklung kooptiert wurden. Andernfalls sollte man wohl bei allen Gruppen dasselbe Pax-Gen erwarten. Die Autoren vermuten darüber hinaus, dass die kooptierten Pax-Gene im gemeinsamen Vorfahren der Cnidarier und Bilaterier bereits überschneidende Aufgaben bei der Augenentwicklung hatten. Damit sehen sie die monophyletische Entstehung aller Augentypen wieder als mögliche und gut begründete Hypothese im Rennen.
 |
| Abb. 2: Modell einer Evolution der Pax-Gene, die für die Entwicklung von Augen rekrutiert wurden. 1: Genverdopplung führt zu verschiedenen Pax-Klassen, 2: Das ursprüngliche Auge entstand und verschiedene Klassen von Pax-Genen wurden redundant für die Augenentwicklung rekrutiert. 3: In allen drei Linien wurde ein spezifisches Pax-Gen für die Augenentwicklung ausgewählt. 4: Pax-Gene, die für die Augenentwicklung verantwortlich sind, werden in einigen Bilateriern (Zweiseitentiere) verändert. Pax-Gene gibt es auch bei augenlosen Schwämmen; man vermutet evolutionstheoretisch, dass sie ursprünglich eine andere Funktion hatten. (Nach Suga et al. 2010) |
|
Bemerkenswert ist die Art und Weise, wie die Autoren die vermuteten Kooptionen beschreiben: Sie sprechen einerseits von einer Wahl unterschiedlicher Pax-Gene in den verschiedenen Tierklassen (S. 14267) und bezeichnen andererseits den hypothetischen Vorgang der Nutzung verschiedener Pax-Gene als „molecular-level opportunism“, den es häufig auch bei den Crystallinen (den Linsen-Proteinen) gegeben habe. Gemeint ist damit, dass in unterschiedlichen Linien zwar ähnliche, aber eben nicht identische Gene für einen bestimmten Zweck rekrutiert wurden. Die neuen Untersuchungen legen diesen Vorgang bei den Pax-Genen nahe. Es gibt aber auch Fälle, bei denen ganz unterschiedliche Gene rekrutiert werden, das ist bei den Linsencrystallinen vielfach der Fall (Piatigorski 2008).
Hinter diesen Beschreibungen stehen aber schwerwiegende Probleme für einen ungerichteten, natürlichen evolutionären Prozess. Schon eine einzige Übernahme eines Gens in ein neues, komplexes Netzwerk von Beziehungen erfordert vielfache Abstimmungen, insbesondere wenn es sich wie bei den Pax-Genen um Steuergene handelt, die eine prominente Position in den Regulationskaskaden einnehmen. Die Erklärungsprobleme potenzieren sich, wenn Kooptionen (wie im Falle von Pax-A, Pax-B und Pax-6) mehrfach unabhängig in ähnlicher Weise zum selben spezifischen Zweck der Strukturentwicklung des Auges erfolgt sein sollen.
|
|
Constraints
 |
| Bei den Bilateria, den Hydrozoa und den Cubozoa werden Pax-Gene aus ver-schiedenen Pax-Unterfamilien verwendet. |
 |
|
An dieser Stelle wird manchmal argumentiert, dass ähnliche Konstellationen bestimmte Kooptionsvorgänge begünstigen könnten; man spricht dann von „Kanalisierung“: Entwicklungszwänge, sog. constraints lenkten die weitere evolutive Entwicklung in eine bestimmte Richtung, wodurch es zu den Konvergenzen komme. Dies mag nachvollziehbar sein, wenn es um Feinabstimmungen und Spezialisierungen geht. Hier geht es aber um eine Neuverschaltung innerhalb eines bereits bestehenden komplexen genetischen Netzwerks. Der Hinweis auf constraints bietet keine Erklärung, da an diesem konkreten Beispiel der Augenentwicklung nicht gezeigt werden kann, worin diese bestehen sollen und wie sich diese auf
genetischer oder morphologischer Ebene darstellen.
Die Annahme von constraints, die eine Kanalisierung und Parallelentwicklung in die Wege leiten, bleibt rein hypothetisch, solange nicht gezeigt wird, wie eine vorausgegangene Konstellation die nachfolgende Kooption begünstigt haben könnte. Wie soll ein Pax-Steuergen, das im Gesamtgefüge der Genwirkungen zunächst irgendeine andere Aufgabe unabhängig vom Sehen hatte, dafür prädestiniert gewesen sein, so dass es mit einer scheinbaren Notwendigkeit als Masterkontrollgen für die Bildung eines Auges kooptiert wurde? Pax-Gene haben verschiedene Funktionen – warum wurden sie mehrfach (d. h. mindestens zweimal oder dreimal) gerade für die Sehfunktion kooptiert? Warum Pax-Gene aus verschiedenen Familien? Da die Pax-Gene an prominenter Stelle wirken, braucht eine Kooption nach allem, was bekannt ist, wie erwähnt viele Abstimmungen. Wie kann dies dreimal unabhängig in unterschiedlichen evolutionären Stammeslinien in ähnlicher Weise mit ähnlichem Ergebnis ohne Planung so erfolgreich ablaufen?
Dank: Dr. Henrik Ullrich danke ich für wertvolle Hinweise.
|
Literatur
- Gehring W (1996)
- The master control gene for morphogenesis and evolution of the eye. Genes Cells 1, 11-15.
- Gregory R (2008)
- The evolution of complex organs. Evo. Edu. Outreach 1, 358-389.
- Halder H, Callaerts P & Gehring WJ (1995)
- Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila. Science 267, 1788-1792.
- Nilsson D-E, Gislén L, Coates MM, Skogh C & Garm A (2005)
- Advanced optics in a jellyfish eye. Nature 435, 201-205.
- Piatigorski J (2008)
- A Genetic Perspective on Eye Evolution: Gene Sharing, Convergence and Parallelism. Evo. Edu. Outreach 1, 403-414.
- Suga H, Tschoppa P, Graziussia DF, Stierwald M, Schmid V & Gehring WJ (2010)
- Flexibly deployed Pax genes in eye development at the early evolution of animals demonstrated by studies on a hydrozoan jellyfish. Proc. Natl. Acad. Sci. 107, 14263-14268.
|
|  |
