 |
|
| |
|
Einem von vielen Rätseln in der Natur scheinen Forscher auf die Schliche gekommen zu sein: die phänomenalen Flugeigenschaften der Ahornfrüchte. Bekanntlich besteht die Frucht des Ahorns aus zwei Teilen, die sich bei der Reife trennen und dadurch zu Teilfrüchten werden. Diese werden in der Botanik meist etwas verkürzt als Früchte, im Volksmund hingegen als Samen bezeichnet. Sie besitzen je einen auffälligen Flügel und umschließen in ihrem unteren, dem Flügel abgewandten Teil jeweils einen Samen. Schon jedes Kind hat seinen Spaß mit der Beobachtung, wie solche Früchte rotierend zu Boden schweben (Abb. 1).
 |
| Abb. 1: Teilfrucht des Ahorns im Propellerflug (Fotomontage) Foto: fotolia.com |
|
Ein wesentlicher, bereits länger bekannter Faktor zur Verzögerung der Fallgeschwindigkeit ist also die schraubenförmige Rotationsbewegung, die jedoch nicht ausschließlich für eine Abbremsung sorgt. In einer kurzen Sturzflugphase nimmt die Frucht zunächst Geschwindigkeit auf. Die Anströmung der Luft führt nun durch die spezielle Beschaffenheit des Flügels zu einer Rotationsbewegung um die Achse der Falllinie des Schwerpunktes des Flugobjekts. Dieser liegt je nach Reifegrad nahe dem Flügelansatz. Der Flügel nimmt in der ersten Flugphase rotationsbedingt eine relativ flache Stellung ein und wirkt nun als Propeller-Tragfläche. Er beginnt als Schraubenflieger der Luft einen größeren, scheibenförmigen Widerstand entgegenzusetzen. Diese „Propellerbewegung“ ist z.B. Vorbild für den Tragschrauber oder Gyrokopter, dessen Propeller sich alleine durch die Anströmgeschwindigkeit der Luft bei Horizontalbeschleunigung in Bewegung setzt und so ohne Motorantrieb für Auftrieb sorgt.
Doch damit nicht genug. Durch die Rotation erfährt der Flügel der Teilfrucht eine Anströmung der Luft gegen die Flügelvorderkante und dies führt zu speziellen Strömungseffekten. In den Ahornfrüchten steckt also noch mehr aerodynamisches Know how, als bisher bekannt war, wie David Lentink und seine Kollegen von der niederländischen Universität Wageningen jetzt in aufwändigen Versuchen herausfanden. Zunächst wurde am 10-fach vergrößerten Kunststoffmodell im Öltank die Bewegung der Frucht beobachtet und es wurden mit Hilfe von kleinsten Glaskügelchen im Laserlicht Strömungslinien sichtbar gemacht. In anschließenden Versuchen mit echten Ahornfrüchten im Windkanal konnte bei Zugabe von Rauch bestätigt werden, dass sich an der Oberseite der Vorderkante des Flügels ein deutlicher Wirbel (leading edge vortex) ausbildet. Es handelt sich quasi um einen Wirbelzopf, der sich vom Schwerpunkt der Frucht spiralförmig nach außen zur Flügelspitze hin bewegt und dort gegen die Flugrichtung abknickt, um sich schließlich aufzulösen. Dadurch entsteht ein zusätzlicher Unterdruck auf der Oberseite des Flügels und erhöht somit dessen Auftrieb. Das Erstaunliche daran ist, dass manche Insekten ebenso wie Kolibris und Fledermäuse durch eine spezielle Kombination aus Auf- und Abschlag sowie einer Rotationsbewegung des Flügels um die Längsachse ähnliche auftriebverstärkende Wirbel erzeugen.
Neben der Möglichkeit, diesen Effekt für moderne Fluggeräte nutzen zu können, wird auch an diesem Beispiel wieder deutlich, dass sich konstruktive Elemente in der Natur oft an stammesgeschichtlich nicht verknüpften Punkten finden, nämlich dort, wo sie von ihrer Funktionalität her sinnvoll sind. Es handelt sich um ein außergewöhnliches Beispiel einer Konvergenz, also einer mehrfach unabhängigen Entstehung. Gleiche komplizierte Erfindungen ohne Erfinder?
[Lentink D et al. (2009) Leading-Edge Vortices Elevate Lift of Autorotating Plant Seeds. Science 324, 1438-1440; Brück J & Kuhn B (2008). Bionik, BIONIK – der Natur abgeschaut. Köln: NGV. Internet: http://www.n-tv.de/wissen/weltall/Der-Flug-der-Ahornsamen...; http://www.faz.net/s/...]
|

|
In der sogenannten Kambrischen Explosion entstanden in einem geologisch sehr kurzen Zeitraum („Augenzwinkern“) praktisch alle heutigen Tierstämme. Aus Sedimentgesteinen vor dem Kambrium sind zwar einige Fossilien bekannt, doch diese eignen sich kaum als Vorläufer für die Tierwelt, die ab dem Kambrium auftritt. Ohne weitere bekannte Vorläufer kommt es zu einer plötzlichen Überfülle an tierischen Bauplänen. Dieses Rätsel möchte eine neue Disziplin, „molekulare Paläobiologie“ erhellen. In einem neuen Artikel stellen Petersen et al. (2009) drei neue Befunde vor, die ihrer Meinung nach Licht in das Dunkel bringen könnten: 1. Vergleicht man die Körperbaupläne der Trilobiten über die geologische Zeit, so ist die Variabilität der frühen Arten höher und sie nimmt im Laufe der Zeit vom Kambrium zum Devon hin ab, bevor die Trilobiten im Perm aussterben. 2. Eine neue Form der Regulation wurde entdeckt: Regulation durch microRNA. MicroRNAs sind etwa 22 Nukleotide lang und sie binden sich an die Genkopien (mRNAs), die normalerweise in Proteine übersetzt werden würden. Durch die Bindung der microRNA an die mRNA kommt es aber zur Blockierung und zum Abbau der betroffenen Genkopien. 3. Im Laufe der Evolution erwarben die Mehrzeller immer mehr solcher regulativer microRNAs.
Am Anfang der Kambrischen Evolution steht also ein Organismus, dessen molekulare Maschinerie aufgrund ihrer Primitivität viele Möglichkeiten eröffnet und den „Raum aller Baupläne“ dadurch effektiv „absuchen“ kann. Das soll das Fenster für die Entstehung der vielen unterschiedlichen Baupläne in der Kambrischen Explosion öffnen. Dann werden vorteilhafte Baupläne mit ihrer noch unpräzisen Genregulation durch die microRNAs präzisiert und damit in eine bestimmte Richtung hin kanalisiert und konserviert. Der Vorteil der microRNA-Regulation liegt darin, dass das betreffende regulierte Gen nicht verloren geht, sondern für spätere Evolutionsschritte weiterhin zur Verfügung steht.
Kritik. Petersen et al. (2009) erklären nicht – wie so häufig – die Entstehung der Anfangszustände. Es gibt nach wie vor keine Antwort auf die Frage: Wie kommt es zu einem Lebewesen mit einer anfangs (hypothetischen!) unpräzisen aber dennoch lebensfähigen Genregulation? Dieses Lebewesen muss komplex genug sein, um die Evolution praktisch aller bekannten Tierstämme zu erlauben. Zum anderen ist der Mechanismus der Genregulation durch microRNAs selbst durchaus komplex. Wie ist dieser entstanden? Die in der nachfolgenden Evolution neu hinzugekommenen microRNAs mussten im passenden Moment hergestellt werden, mehrere Enzymkomplexe sind daran beteiligt und die blockierten mRNAs mussten abgebaut werden. Wie entstand dieser komplexe Mechanismus? Weitergehende Fragen der Evolvierbarkeit solcher Systeme sind davon noch völlig unberührt.
[Peterson KJ, Dietrich MR & McPeek MA (2009) MicroRNAs and metazoan macroevolution: insights into canalization, complexity, and the Cambrian explosion. Bioessays 31, 736-747.]
|
|
Ihre Stabilität und Größe verdanken unsere Bäume einem besonderen Stoff: Lignin (lat. lignum, Holz). Dabei handelt es sich um komplexe aromatische chemische Verbindungen, die aus verschiedenen Einzelbausteinen zusammengesetzt sind (Heteropolymere) und in die sekundäre pflanzliche Zellwand eingelagert werden (Verholzung oder Lignifizierung). Dort bilden Lignine eine dichte Matrix, die an Cellulosefasern bindet und andere Bestandteile der Zellwand miteinander verknüpft. Durch diese Einlagerung bewirkt Lignin eine Verholzung der Zelle und ermöglicht deren Festigkeit. Die Trockenmasse verholzter Pflanzen besteht zu 20% bis 30% aus Lignin. Evolutionstheoretisch werden lignifizierte Zellwände als Schlüsselneuheiten bei der Entstehung von Landpflanzen interpretiert.
Nun berichten Mortone et al. (2009) von der Entdeckung sekundärer Zellwände mit Lignin in der Rotalge Calliarthron cheilosporioides, die in der Gezeitenzone lebt. Bisher waren derart spezialisierte Zellwände und Lignine nur von Gefäßpflanzen bekannt. Da die Rotalgen in keiner näheren Verwandtschaftsbeziehung zu den Gefäßpflanzen stehen, drängt sich die Annahme einer konvergenten (zweimaligen unabhängigen Entstehung) von Lignin auf. Evolutionstheoretisch ist dies angesichts der Komplexität dieser Moleküle sehr problematisch, da deren biochemische Synthese ausgesprochen komplex ist (Mortone et al. 2009, 170). Welche andere Möglichkeit gibt es, wenn Evolution zugrundegelegt wird? Die Autoren vermuten gemeinsame Synthesewege bei gemeinsamen Vorfahren von Rotalgen und Gefäßpflanzen, so dass man es doch wenigstens zum Teil mit sogenannten „tiefen Homologien“ zu tun hätte. („Homolog“ werden Ähnlichkeiten genannt, die auf gemeinsame Vorfahren zurückgeführt werden.) Das würde die Konvergenz mindestens weniger ausgeprägt erscheinen lassen. Ein Beispiel eines solchen Synthesewegs wurde tatsächlich gefunden. Damit ist aber nur ein möglicher Baustein für eine unabhängige Lignin-Synthese nachgewiesen. Die Frage, weshalb und wie schon vorhandene Synthesewege genutzt wurden, um denselben chemischen Stoff zu synthetisieren, bleibt dennoch offen. Mortone et al. (2009, 172) „spekulieren“, dass die Lignin-Biosynthese-Wege im evolutionstheoretisch anzunehmenden gemeinsamen Vorfahren von Rot- und Grünalgen (von Letzteren sollen die Gefäßpflanzen abstammen) die Aufgabe hatten, die Zellen vor mikrobiellen Infektionen und UV-Strahlung zu schützen. Aus einer Schöpfungsperspektive ist das unabhängige Auftreten komplexer Merkmale im Sinne eines „Baukastensystems“ zu verstehen, aus dem sozusagen nach Bedarf funktionell passende Teile entnommen werden können.
[Mortone PT, Estevez JM, Lu F, Ruel K, Denny MW, Somerville C & Ralph J (2009) Discovery of lignin in seaweed reveals convergent evolution of cell-wall architecture. Curr. Biol. 19, 169-175.]
|
 |
| Abb. 1: Vogelartige Fußspuren aus der Santo Domingo-Formation der Obertrias. Balken: 10 cm. (© Ricardo N. Melchor, Abdruck mit freundlicher Genehmigung) |
|
Im Jahr 2002 berichteten Melchor et al. über hunderte fossiler Fußabdrücke aus Argentinien, die bezüglich Abfolge und Morphologie verblüffend vogelähnlich aussehen (vgl. Studium Integrale Journal 9, 95-96; Abb. 1). Sie werden der Spurengattung (Ichnotaxon) Gruipeda zugeordnet. (Von Ichnotaxa kennt man nur Spuren, nicht aber deren Verursacher.) Diese Fußabdrücke sind allerdings 55 Millionen Jahre älter als die ältesten Skelettfunde früher Vögel wie z.B. des bekannten Archaeopteryx und werden erdgeschichtlich in den Übergang Obertrias/Unterjura gestellt. In einer umfangreichen Studie wurden diese Spuren nun mit verschiedenen Arten von Fußspuren heutiger Vögel verglichen, die in küstennahen Biotopen – Tümpeln mit zeitweise trockenfallenden Ufern – vorkommen, weil sie den fossilen Lebensräumen mutmaßlich gleichen oder ähneln (Genise et al. 2009). Als moderne Vergleichsgruppe wurden Watvögel (Strandläufer Calidris sp.) ausgewählt. Den unterschiedlich ausgeprägten Fußspuren der Watvögel ließen sich bestimmte Verhaltensweisen zuordnen, z.B. vor- und seitwärts gerichtete Laufbewegungen, Stochern nach Nahrung oder Flugbewegungen. Fünf dieser Verhaltensweisen konnten auch auf die fossilen Spuren bezogen werden, darunter Merkmale, die auf Flugfähigkeit der Verursacher schließen lassen. Die Autoren werten dies als deutlichen Hinweis auf eine Vogelverwandtschaft der Verursacher der Spuren. Alternativ wäre denkbar, dass sich entsprechende Füße und die von ihnen gesetzten Arten der Spuren konvergent bei zweibeinigen Theropoden-Dinosauriern entwickelt haben, doch ist diese Annahme spekulativ. Das Alter der Schichten, das mittels fossiler Bäume, radiometrischer Datierungen und paläomagnetischer Studien bestimmt wurde, wird von den Autoren aufgrund dieser Funde angezweifelt. Damit der Befund besser mit der zeitlichen Einordnung von Körperfossilien zusammenpasst, müssten die betreffenden Schichten jünger sein (Genise et al. 2009, 143, 159, 160). Weitere vogelähnliche Fußspuren waren schon früher in Südafrika entdeckt worden, ebenfalls im Bereich der Grenze Obertrias/Unterjura.
Das Hinterfragen eines (auch) radiometrisch bestimmten Alters aufgrund unerwarteter Fossilfunde ist ungewöhnlich. In einem Kurzzeitszenario wäre das deutlich frühere Auftreten von Spuren vor der Überlieferung von Körperfossilien nicht so ungewöhnlich (vgl. Stephan 2002). Denn ein Individuum kann sehr viele Spuren hinterlassen, aber nur einen Körper. Dass Spurenfossilien vor Körperfossilien gefunden werden, ist so gesehen nicht überraschend, wohl aber dass beides durch einen Zeitraum von 55 Millionen Jahren getrennt sein soll. Kein Wunder also, wenn die zeitliche Einstufung der Spurenfossilien in Frage gestellt wird.
[Genise JF, Melchor RN, Archangelsky M, Bala LO, Straneck R & de Valais S (2009) Application of neoichnological studies to behavioural and taphonomic interpretation of fossil bird-like tracks from lacustrine settings: The Late Triassic–Early Jurassic? Santo Domingo Formation, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 272, 143-161; Melchor RM, De Valais S & Genise JF (2002) Bird-like fossil footprints from the late Triassic. Nature 417, 936-938; Stephan M (2002) Der Mensch und die geologische Zeittafel. Holzgerlingen; s. auch: http://www.arn.org/blogs/...did_birds_fly_in_the_late_triassic]
|
|
In den letzten Jahren galt die Hypothese, dass zweibeinige kleine Raubdinosaurier (Theropoden) die evolutionären Vorfahren der Vögel seien, als zunehmend gesichert. Überspitzt formuliert lautet das Schlagwort: „Vögel sind überlebende Dinosaurier.“ Nur wenige Paläontologen und Ornithologen widersprachen.
Als Kritikpunkte an der Dino-Vogel-Hypothese wurden genannt (vgl. Junker 2005): 1. Die den Vögeln ähnlichsten Theropoden stammen alle aus der Oberkreide und sind damit sehr viel jünger als der oberjurassische „Urvogel“ Archaeopteryx. 2. Fast alle Theropoden sind zweibeinige Läufer und wegen ihres zu großen Gewichts aus aerodynamischen Gründen nicht als Vogelvorläufer geeignet. 3. Einem hypothetischen evolutiven Umbau von Theropoden zum Vogel-Bauplan stehen konstruktive Aspekte entgegen: So entspricht die Vogelhand den Fingern II, III und IV, während die Theropoden die Finger I, II und III besaßen. Eine homeotische Transformation als Erklärung für diese Verschiebung ist hypothetisch und wegen eines fehlenden Selektionsdrucks schwer zu begründen (vgl. Ullrich 2008). Ein kontrovers diskutiertes weiteres Beispiel ist der Bau der Lunge. Fossilfunde deuten darauf hin, dass Theropoden eine krokodilartige Lunge besaßen, die nach Auffassung einiger Forscher konstruktiv nicht in die ganz anders gebaute Vogellunge umgebaut werden könne (Jones & Ruben 2001).
Der letztgenannte Kritikpunkt erhält neuerdings durch eine Studie von Quick & Ruben (2009) weitere Unterstützung. Die beiden Forscher weisen auf einen bisher offenbar nicht beachteten anatomischen Zusammenhang hin: Lungenfunktion und Oberschenkelanatomie hängen bei Vögeln eng miteinander zusammen. Damit hat es folgende Bewandtnis: Der Oberschenkelknochen sitzt bei den Vögeln relativ fest, und die Hauptbewegung findet im Unterschenkel und im Fuß statt. Diese Anatomie hängt damit zusammen, dass Knochen und Muskulatur der Oberschenkelregion benötigt werden, um die Luftsack-ähnliche Lunge der Vögel vor dem Kollabieren zu bewahren. Die Position der Oberschenkelknochen und Muskeln ist also entscheidend für die Lungenfunktion der Vögel, und erst diese gibt den Vögeln ausreichend Lungenkapazität für den Flug. Die einzigartige Lungenstruktur wird gebraucht, weil die Vögel etwa 20 Mal mehr Sauerstoff als Reptilien benötigen.
Die Theropoden-Dinosaurier weisen dagegen eine ganz andere Anatomie der beweglichen Oberschenkelknochen auf, die nicht in die Vogelanatomie überführbar sei. Daher müsse die Evolution der Vögel unabhängig von den Dinosauriern verlaufen sein. Quick & Ruben stellen damit eine etablierte Sicht in Frage und vermuten gemeinsame Vorfahren von Dinosauriern und Vögeln unter den älteren thecodonten Reptilien (Urwurzelzähnern). Sie weisen auch auf das oben genannte Problem hin, dass die Dinosaurier, von denen die Vögel abstammen sollen, durchweg in erheblich jüngeren Schichten als die ältesten Vögel gefunden seien.
Bemerkenswert an der Position von Quick & Ruben ist, dass sie behaupten, eine konstruktive Umwandlung sei unmöglich. Dann stellt sich aber die Frage, ob diese Unmöglichkeit auch anderen postulierten Übergängen entgegensteht. Die Atmung bei Vögeln ist auf so vielfältige Weise mit anatomischen Konstruktionen verwoben, dass der Umbau zur Vogellunge ein entsprechend komplexer Vorgang gewesen sein muss. Ob natürliche Evolutionsmechanismen das erlauben, kann ernsthaft bezweifelt werden. Für evolutionstheoretische Deutungen sind Merkmalsübereinstimmungen – einer der Belege für die Dino-Vogel-Hypothese – zu wenig, um evolutive Übergänge zu begründen. Man kann die Mechanismenfrage nicht ausblenden, zumal Merkmalsübereinstimmungen auch funktionelle Gründe haben können, ohne dass ein Bezug auf Evolution genommen werden müsste.
[Jones TD & Ruben JA (2001) Respiratory structure and function in theropod dinosaurs and some related taxa. In: Gauthier J & Gall LF (eds) New Perspectives on the Origin and Early Evolution of Birds. New Haven, pp 443-461; Junker R (2005) Der Ursprung der Vögel – ein Update. Stud. Int. J. 12, 51-57; Quick DE & Ruben JA (2009) Cardio-pulmonary anatomy in theropod dinosaurs: implications from extant archosaurs. J. Morphol., 2009, May 20 (online); Ullrich H (2008) Sind Vogelflügel umgestaltete Dinosaurierhände? Zum Konflikt zwischen fossilen und entwicklungsbiologischen Daten bei der phylogenetischen Herleitung des Vogelflügels. Stud. Int. J. 15, 18-30; Vögel doch nicht Nachfahren der Dinos? http://www.scinexx.de/wissen-aktuell-10026-2009-06-10.html]
|
|
Die Hypothese der Abstammung der Vögel von zweibeinig sich fortbewegenden kleinen Raubdinosauriern (vgl. das vorige Streiflicht) wurde in den letzten Jahren vor allem auch durch zahlreiche Funde von Dinosauriern postuliert, die federartige Strukturen besitzen. Allerdings ist die Federnatur dieser Hautstrukturen teilweise umstritten (Feduccia et al. 2005). Zheng et al. (2009) berichten nun vom Fund eines Dinosaurier-Teilskeletts mit Schädelresten, genanntTianyulong confuciusi. Es gehört zur Untergruppe der Ornithischia und besitzt unverzweigte filamentöse (fadenförmige) Hautstrukturen, die als Protofedern interpretiert werden. Die bis zu 60 mm langen und 0,4 mm dicken Filamente wurden an drei Stellen in der Nähe der Wirbelsäule gefunden und scheinen hohl gewesen zu sein. Die Autoren interpretieren dies als äußere Hautstrukturen; es soll sich nicht um innere Versteifungselemente der Haut handeln.
Besonders bemerkenswert ist bei diesem Fund, dass er nicht der Dinosaurier-Großgruppe Saurischia (Echsenbeckensaurier) mit der Untergruppe Theropoda angehört, aus der die Vögel hergeleitet werden, sondern der anderen Großgruppe, den Ornithischia (Vogelbeckensaurier). Sie sollen sich schon sehr früh (in der Mittel-/Obertrias) von den Saurischiern getrennt haben. Damit müssten sich die Hautstrukturen von Tianyulong unabhängig von den Hautstrukturen der Saurischier entwickelt haben. Wellnhofer (2009) ist der Auffassung, der letzte Beweis fehle, dass es sich wirklich um epidermale und nicht um innere Strukturen handle. Dazu müsse sicher geklärt werden, ob sie wirklich hohl waren, was für eine äußere Hautstruktur sprechen würde. Gegen eine Festigungsfunktion führt er an, dass die Filamente im Schwanzbereich extrem lang sind und die Schwanzwirbelsäule ohnehin durch verknöcherte Sehnen versteift gewesen sei. Vergleichbare epidermale Bildungen kennt man schon lange von den ebenfalls nicht näher verwandten Flugsauriern. Der Besitz haarartiger Körperanhänge steht in diesen Fällen offenbar nicht im Zusammenhang mit der Entstehung von Federn, und ob das bei den Gattungen der Fall ist, die man aus anatomischen Gründen eher als Vogelvorläufer ansieht, ist ebenso fraglich. Ein Problem für die Deutung dieser Strukturen ist die Unklarheit ihrer Funktion.
Wie die bisher bekannten Dinosaurier mit federähnlichen Strukturen ist auch Tianyulong jünger als der Urvogel Archaeopteryx, der echte, „moderne“ Federn besaß. Die Gruppe der Heterodontosauridae, zu der Tianyulong gehört, ist aber schon aus dem frühen Jura bekannt (Alter 200 Millionen Jahre nach herkömmlicher Zeitrechnung), so dass Tianyulong in der Unterkreide (vor 125-130 Millionen Jahren) ein lebendes Fossil darstellt. Die systematische Stellung der Heterodontosauridae ist unsicher, sie gelten aber als eine der ursprünglichsten Gruppen der Ornithischia (Zheng et al. 2009).
[Feduccia A, Lingham-Soliar T & Hinchliffe JR (2005) Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological Evidence. J. Morphol. 266, 125-166; Wellnhofer P (2009) Protofedern bei Vogelbeckensauriern? Nat. Rdsch. 62, 311-312; Zheng XT, You HL, Xu X & Dong ZM (2009) An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures. Nature 458, 333-336.]
|
|
Eine der zentralen Neuheiten der Vögel gegenüber ihren mutmaßlichen Vorläufern nach der Evolutionslehre sind die Federn. Federn können verschiedene Funktionen ausüben, sind aber nur dann für den Flug brauchbar, wenn sie entsprechend detailliert gebaut, zweckmäßig und flexibel in der Körperoberfläche verankert und in einem funktionalen Federkleid zusammengefügt sind. Außerdem benötigt ein Vogel ein entsprechendes Verhaltensrepertoire, um den Flugapparat mit allen seinen Einzelteilen überhaupt nutzen zu können. Es ist klar, dass der Sprung von federlosen Reptilien zu flugfähigen Vögeln so groß ist, dass er evolutiv allenfalls dann überbrückt werden kann, wenn es Zwischenstufen mit anderen Funktionen gibt. So wird beispielsweise angenommen, dass einfachere Federstrukturen – sogenannte Protofedern – ursprünglich der Wärmeisolierung dienten, zumal es auch heute Federn gibt, die nur diese Funktion ausüben.
Eine notwendige, wenn auch bei weitem nicht hinreichende Voraussetzung für die Plausibilität einer evolutiven Entstehung der Federn ist der Nachweis verschieden ausgeprägter Federstrukturen. Hierzu existieren zum einen theoretische Modelle (Prum 1999, Prum & Brush 2003), die in den vergangenen Jahren zum anderen mit Funden mutmaßlicher Protofedern gestützt wurden (kritisch dazu aber Peters 2001). Ein Fund von einfachen federähnlichen Strukturen in Bernstein aus der oberen Unterkreide Frankreichs scheint hier einen wichtigen Baustein zu liefern (Perrichot et al. 2008), da er möglicherweise zwischen zwei Stufen des Modells von Prum vermittelt. Es handelt sich um sieben identische Federn, die Seite an Seite im Bernsteinstück liegen (Abb. 1). Ihre Basis scheint zu fehlen. Die drei am besten untersuchbaren Federn sind 2,3, 1,6 und 1,1 mm lang, also winzig, und fiederig verzweigt. Die Rachis (Schaft) ist abgeflacht. Den Federästen fehlen die Federstrahlen, die die Äste der Federn bei vielen heutigen Vögeln miteinander verknüpfen. Ihre Form kommt Daunenfedern, Schmuck- oder Doppelfedern (Feder mit zwei Schäften, an der Basis verzweigt) morphologisch am nächsten; diese sind jedoch den Konturfedern nicht ähnlich. Anders als typische Daunenfedern haben sie eine dicke und lange Rachis, deren Äste allgemein von der sehr kurzen Spitze der Rachis abgehen (Perrichot et al. 2008, 1198). Die Autoren betrachten den Fund als intermediäre Ausprägung in der hypothetischen Federevolution, deren Existenz durch Theorien vorhergesagt worden sei, die sich an der ontogenetischen Federentwicklung orientieren (Prum 1999).
Ob es sich um eine evolutive Übergangsform handelt, ist aber aus mehreren Gründen fraglich. Da die fossilen Federn morphologisch Daunenfedern stark ähneln, könnte es sich auch um solche Federn handeln, auch wenn eine Entsprechung mit heutigen Daunenfedern nicht durchweg vorliegt. Da die Fossilien so klein sind, müssten sie mit Daunenfedern frisch geschlüpfter Küken verglichen werden, nicht mit denen ausgewachsener Vögel. Es könnte sich auch um degenerierte Federn handeln. Federn mit einem Schaft, aber ohne Federstrahlen an den Ästen, sind von heutigen Vögeln wie den Kiwis bekannt. Wie alle anderen bisher entdeckten mutmaßlichen Protofedern ist auch der neue Bernsteinfund deutlich zu jung für die Deutung als Vorfeder, denn die den heutigen Federn vergleichbaren Archaeopteryxfedern sind älter. Unklar ist auch, welche Selektionsvorteile die Ausprägung der fossilen Bernsteinfedern gehabt haben könnten.
[Anonymus: Annotations from the literature. http://www.grisda.org/origins/62026.pdf; Perrichot V, Marion L, Neraudeau D, Vullo R & Tafforeau P (2008) The early evolution of feathers: fossil evidence from Cretaceous amber of France. Proc. R. Soc. 275B, 1197-1202; Peters DS (2001) Probleme der frühen Vogelevolution. I. Die Sache mit den Federn. Nat. Mus. 131, 387-401; Prum RO (1999) Development and evolutionary origin of feathers. J. Exp. Zool. 285B, 291-306; Prum RO & Brush AH (2003) Zuerst kam die Feder. Spektr. Wiss. 10/2003, 32-41; Tyler D (2008) Feathers with „remarkably primitive features“. http://www.arn.org/blogs/index.php/literature/2008/02/26/feathers_with_remarkably_primitive_featu]
|
|
In einer kürzlich veröffentlichten Studie berichten Dussutour und Mitarbeiter über eine Anpassung der Blattschneiderameisen an äußere Gegebenheiten, die bisher so nicht beobachtet worden war. Die Ameisen schneiden Blätter und transportieren die Blattstücke in ihr Nest, um darauf Pilze zu züchten. Die Forscher studierten nun das Verhalten der Ameisen, nachdem sie die lichte Höhe des Transportwegs auf 1 cm Höhe verringerten. Nach kurzer Zeit schnitten die Ameisen kleinere und rundere Blattfragmente. Um nun die kleinere Blattmenge pro Transportameise auszugleichen, wurden nicht mehr Ameisen ausgeschickt als vorher, sondern mehr Ameisen kehrten beladen heim. Dadurch wurde die Transportmenge an Pflanzenmaterial sogar überkompensiert, d.h. mit Verengung trugen die Ameisen mehr ein als ohne sie. Woher wissen die Ameisen nun, dass sie kleinere Blattstücke schneiden müssen, um den Tunnel passieren zu können? Entweder können Ameisen, die nach draußen laufen, bereits beim Durchqueren des Tunnels dessen lichte Höhe abschätzen (solch ein Verhalten konnte bei Blattschneiderbienen beobachtet werden) oder sie gehen nach Versuch und Irrtum vor, d.h. sie merken sich nach einiger Zeit, mit welcher Blattgröße sie zurücklaufen können und mit welcher nicht. Welche der beiden Vermutungen die richtige ist, ist noch völlig offen. Insgesamt reagieren die Ameisen auf zwei Ebenen, um der Situation zu begegnen. Einerseits auf der Ebene des Individuums: „Schneide kleinere Blattstücke!“, und andererseits auf Ebene der Gemeinschaft: „Hilf beim Transport!“. Dussutour kommentiert die Ergebnisse in einem Interview mit den Worten: „Es legt nahe, dass die Tiere viel klüger sind, als wir dachten.“
[Dussutour A, Deneubourg J-L, Beshers S & Fourcassié V (2009) Individual and collective problem-solving in a foraging context in the leaf-cutting ant Atta colombica. Anim. Cogn. 12:21–30; Boeddeker M (2008) Logistik, die mitdenkt. http://www.wissenschaft.de/wissenschaft/news/292611.html]
|
|
Im Rahmen evolutionärer Erklärungsansätze werden auch Theorien zur Evolution der Nahrungssuche in Räuber-Beute-Systemen entwickelt. Anhand von Mikroorganismen wurde jetzt der Einfluss von viel oder wenig Beute auf das Jagdverhalten der Räuber untersucht. Dabei konnte gezeigt werden, dass die Suchgeschwindigkeit stark durch die Verteilung der Beute geprägt wird. Es wurde aber auch deutlich, dass ein effektiveres Jagdverhalten mit einer verringerten Überlebensfähigkeit unter ungünstigen Lebensbedingungen einhergeht. Die Optimierung grundlegender Verhaltensweisen ist also begleitet von Nachteilen, die den Fortbestand einer Population in Frage stellen können.
Mikroorganismen zeigen charakteristische Eigenschaften, wie z. B. kurze Generationszeit, geringer Platzbedarf und leichte Handhabbarkeit, die sie als Studienobjekte im Labor attraktiv machen. Für experimentelle Untersuchungen zur Evolution des Jagdverhaltens haben Hillesland et al. (2009) dem räuberisch lebenden Bodenbakterium Myxococcus xanthus in verschiedenen Versuchen kleine Kolonien des Darmbakteriums Escherichia coli als Beute angeboten. Myxobakterien bewegen sich auf Oberflächen (z.B. Bodenpartikel oder Agarplatten) auf unterschiedliche Weise und suchen dabei nach Beute. Treffen sie auf geeignete Mikroorganismen, so setzten sie verschiedene Enzyme und Antibiotika frei, mit denen die Zellen der Beute abgetötet und aufgelöst werden. Die dadurch zugänglichen Proteine und Peptide nutzen die Räuber als Nahrungsmittel. Ist eine Nahrungsquelle erschöpft und die Lebensbedingungen verschlechtern sich, dann bildet M. xanthus durch koordinierte Abläufe vieler einzelner Zellen einen Fruchtkörper, in dessen Inneren sich einzelne Zellen in Sporen differenzieren. Diese sind von dicken Zellwänden umgeben, die sie gegen schädliche Umwelteinflüsse wie beispielsweise Wassermangel und UV-Strahlung schützen und das Überleben auch über längere Perioden unter extremen Mangelbedingungen ermöglichen.
Für die Experimente wurden Myxobakterien fast 1 Jahr lang kultiviert, entweder in Gegenwart von viel Beute (hohe Dichte an kleinen E. coli-Kolonien in der Umgebung) oder bei wenig Beute (geringe Dichte). Die Ausgangskultur von M. xanthus hatte mehr E. coli als einzige Nahrungsquelle zur Verfügung als die Kulturen in beiden Versuchsreihen. In den Versuchen wurden im Labor in quadratischen Petrischalen (12 x 12 cm) mit Agar kleine, nicht mobile E. coli-Kolonien an Gitterpunkten im Abstand von 1 cm (hohe Dichte der Beute) oder 2 cm (geringe Dichte der Beute) platziert. Auf einem Gitterpunkt im Zentrum der Platte wurden die Myxobakterien aufgetragen. Diese schwärmen auf der Suche nach Beute über die Oberfläche der Agarplatte – etwa kreisförmig um die Ausgangsposition – und vermehren sich währenddessen durch Zellteilung (vegetativ). Nach jeweils 14 Tagen wurden schwärmende M. xanthus-Proben zum Impfen neuer Agarplatten (mit hoher bzw. geringer Beutedichte) verwendet. Nach 24 solcher Zyklen wurden die so gezüchteten Myxobakterien bezüglich verschiedener Aspekte ihres Jagdverhaltens mit der Ausgangskultur verglichen. Es zeigte sich, dass bei Selektion auf möglichst schnelle Beutesuche in weniger als 300 Generationen die Geschwindigkeit des Schwärmens bei M. xanthus bis zu 10 mal schneller wurde im Vergleich zur Vorläuferkultur. Myxobakterien, die sich unter geringen Beutedichten ernähren mussten, waren dabei wesentlich schneller als diejenigen, die einen reicher gedeckten Tisch vorfanden.
Die Untersuchungen bestätigen allerdings auch einen bereits beschriebenen genetischen Zusammenhang zwischen Jagdverhalten und der Fähigkeit Fruchtkörper zu bilden. Unter den gewählten Selektionsbedingungen werden Organismen begünstigt, die nicht hohe Zelldichten anstreben, sondern die immer weiter schwärmen und mit möglichst hoher Effizienz neue Nahrungsquellen – sprich Beute – aufspüren. Das bedeutet: Die Bereitschaft und Fähigkeit zur Kooperation wird reduziert. Somit ist die Erhöhung der Jagdgeschwindigkeit (Optimierungsprozess) erkauft mit der verminderten Fähigkeit zur Bildung von Fruchtkörpern. Die Untersuchung von Hillesland et al. (2009) belegt damit einen interessanten Fall von Optimierung von plastischen Eigenschaften bei Mikroorganismen unter bestimmten Selektionsbedingungen. Die Optimierung der Schnelligkeit bei der Jagd in diesem Räuber-Beute System fordert jedoch einen hohen Preis, der unter Umständen den Fortbestand der Kultur gefährdet.
[Hillesland K, Velicer GJ & Lenski RE (2009) Experimental evolution of a microbial predator’s ability to find prey. Proc. R. Soc. 276B, 459-467.]
|
 |
| Abb. 1: Papilio xuthus, ein Verwandlungskünstler, links als Vogelkot getarnt. (Foto: Haruhiko Fujiwara, Abdruck mit freundlicher Genehmigung) |
|
Tarnung im Tierreich ist ein immer wieder spannendes Thema. Beliebt ist das Nachahmen von Pflanzenteilen wie Ästen oder Blättern. Eine sehr ausgefallene Art der Tarnung zeigen die Larven des japanischen Schwalbenschwanzes Papilio xuthus. Sie sehen in den ersten vier Phasen des Raupenstadiums wie Vogelkot aus, schwarz-weiß und klebrig, wie die Ausscheidungen eines Vogels (Abb. 1). Erst wenn die Raupen so groß sind, dass diese Tarnung ihre Überzeugungskraft verliert, färbt sie sich grün wie die Blätter, auf denen sie lebt; sie sieht dann aus wie ein hellgrünes, gerolltes Blatt.
Damit kann sich die Larve des japanischen Schwalbenschwanzes nicht nur genial tarnen, sondern ist auch noch eine regelrechte Verwandlungskünstlerin. An den Metamorphosen (Verwandlungen) ist das Juvenilhormon beteiligt. Vor der Wandlung zum Blattimitat nimmt seine Konzentration stark ab (Futahashi & Fujiwara 2008). Durch die Zugabe eines Hormonersatzes blieb die Vogelkot-Tarnung teilweise bestehen. Beispiele wie diese drängen in besonderem Maße die Frage auf, woher die Programme für solche Formen, Farben und das dazugehörende Timing kommen.
[Futahashi R & Fujiwara H (2008) Juvenile hormone regulates butterfly larval pattern switches. Science 319, 1061.]
|
|
Aus Sedimentsystemen des Jura in Throwbridge (Wiltshire, Großbritanien) sind seit langem verschiedenste sehr gut erhaltene Fossilien bekannt (oft mit Weichteilerhaltung). Nun berichtet die BBC unter Berufung auf Phil Wilby, einen Paläontologen der Britisch Geological Survey von dem außerordentlichen Fund eines fossilen Tintenfischs (Belemnotheutis antiquus, Coeloidea). Bei der Präparation des Fossils wurde ein ca. 2,5 cm langer schwarz gefärbter Tintenbeutel sichtbar. Daraus konnten die Wissenschaftler mit Ammoniaklösung schreibfähige Tinte extrahieren. Eine Skizze des fossilen Tintenfischs und sein Name wurden mit dieser Tinte angefertigt. Phil Wilby wird von BBC mit der Aussage zitiert: „Es ist schwer vorstellbar, dass so etwas wie ein Tintenbeutel – so weich und lappig – in einem Stein der 150 Millionen Jahre alt ist dreidimensional und immer noch schwarz fossil erhalten bleibt.“ („It is difficult to imagine how you can have something as soft and sloppy as an ink sac inside a rock that is 150 million years old.“)
Die Tinte von heutigen Tintenfischen ist ein komplexes Gemisch mit Melanin, Proteinen, Oligosaccchariden und anderen Verbindungen. Wie diese organischen Substanzen über diese lange Zeit erhalten bleiben, so dass eine schwarze Tinte erhalten bleibt, ist bisher ein Rätsel. Wilby spricht von einem Medusa-Effekt: Organismen werden innerhalb von Tagen in Stein umgewandelt. Eine Probe des Tintenbeutels wird derzeit chemisch analysiert. Auf die Ergebnisse darf man gespannt sein, vielleicht eröffnen sie Wege zu einem erweiterten Verständnis von Fossilisationsprozessen.
[BBC NEWS (2009/08/19) http://news.bbc.co.uk/go/pr/fr/-/2/hi/uk_news/england/wiltshire/8208838.stm ; The Times (2009/08/19); http://www.timesonline.co.uk/tol/news/uk/science/article6800939.ece]
|
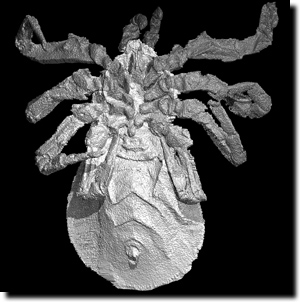 |
| Abb. 1: Cryptomartus hindi, eine der in Siderit erhaltenen Spinnenfossilien aus dem Karbon. Abdruck mit freundlicher Genehmigung des Natural History Museum, London and Imperial College, London. |
|
Seit einigen Jahren werden hochauflösende Röntgentechniken zur zerstörungsfreien Darstellung von Fossilien genutzt. Penney hat zusammen mit Kollegen (2007) Röntgen-Computer-Tomographie zur detaillierten Untersuchung einer in Bernstein erhaltenen kleinen Spinne (ca. 1 mm) genutzt. Bei Paris wurde aus dem frühen Eozän (radiometrisch ca. 53 Millionen Jahre) Bernstein geborgen, der einen Vertreter von Micropholcommatideae, Centrixtricella simoni, als Einschluss enthielt. Die Autoren konnten kleinste Details bei Körpermerkmalen der Spinne dokumentieren und darüber hinaus auch das Innere der Spinne abbilden und so Aussagen über den Zustand der Organe machen. Die Familie der Micropholcommatidae gehört zu den echten Webspinnen und derzeit sind davon 33 Arten beschrieben.
Jetzt haben Garwood et al. (2009) mit ähnlichen Methoden noch sehr viel ältere Spinnentiere beschrieben, die als Fossilien in Siderit – einem Eisencarbonat-Mineral – erhalten sind. Siderit tritt oft vergesellschaftet mit Kohle auf. In der Publikation beschreiben die Autoren zwei Fossilien aus dem Karbon (Duckmantian; radiometrisch ca. 311 Millionen Jahre). Diese bereits früher beschriebenen Fossilien konnten nun mit bisher unbekannten Details beschrieben und dargestellt werden (Abb. 1). Die Fossilien gehören zu den Trigonotarbida, einer ausgestorbenen Gruppe der Spinnentiere (Arachnidae), Vertreter sind vom späten Silur bis ins frühe Perm beschrieben. Die Trigonotarbida weisen viele Ähnlichkeiten mit den Spinnen auf, haben aber keine Spinnwarzen und konnten somit vermutlich keine Spinnseide produzieren.
Die neuen Untersuchungsmethoden eröffnen einen Zugang zu Fossilien mit einer sehr hohen Auflösung, d.h. auch kleinste Details können sichtbar gemacht werden. Dies lässt für die Zukunft noch spannende Einsichten und Erkenntnisse in die Geschichte der Lebewesen erwarten.
[Garwood R, Dunlop JA & Sutton MD (2009) High-fidelity X-ray micro-tomography reconstruction of siderite-hosted Carboniferous arachnids. Biol. Lett. doi: 10.1098/rsbl. 2009.0464; Penney D, Dierick M, Cnudde V, Masschaele B, Vlassenbroeck J, Hoorebeke van L & Jakobs P (2007) First fossil Micropholcommatidae (Araneae), imaged in Eocene Paris amber using X-ray computed tomography. Zootaxa 1623, 47-53.]
|
|
Fische sind in vielen Regionen der Erde eine wichtige Ernährungsgrundlage für den Menschen. Mit der Perfektionierung von Fangmethoden wurde die Überfischung allerdings ab der 2. Hälfte des 20. Jahrhunderts zum Problem: 1852 wurden in der Ostsee etwa 1,26 Millionen Tonnen Kabeljau (Gadus morhua) gefangen gegenüber nur noch 650 000 Tonnen in den frühen 1980er Jahren und 50 000 Tonnen im Jahr 2005. Im Jahr 1993 war der Tiefstand der letzten 15 Jahre, bevor 2003 ein relativer Hochstand erreicht wurde.
Die Überfischung erreichte mittlerweile ein Ausmaß, das sogar das Aussterben des Kabeljaus in der Ostsee möglich erscheinen lässt: Bereits vor 40 000 Jahren (nach konventioneller Zeitrechnung) lebende „moderne“ Menschen (Homo sapiens sapiens) verzehrten neben Fleisch regelmäßig Fisch (Hua et al.). Limburg et al. (2008) verglichen die heutigen Fänge hinsichtlich Größe und Alter beim Fang mit Fängen in der Jungsteinzeit (2500 v. Christus) anhand der Fischabfälle der Siedlung Ajvide an der Südwestküste von Gotland. Die Größe der Fische aus der Jungsteinzeit kann man anhand ihrer Otolithen („Ohrsteine“) bestimmen. Dabei handelt es sich um Kalkgebilde im Gleichgewichtsorgan des Innenohrs. Sie helfen bei der Wahrnehmung von Gravitations- und Zentrifugalkräften und damit bei der räumlichen Orientierung. Wie die knöchernen Wirbelkörper bilden die Otolithen Jahresringe, an denen sich Alter und – wie auch aus den Wirbelkörpern – die Größe eines Fisches ablesen lässt. In der Jungsteinzeit gefangene Kabeljaus waren demnach signifikant größer als heutige.
Wie kam es zu dieser Veränderung? Die Steinzeitmenschen besaßen nicht die technischen Möglichkeiten, große, einen Meter und mehr lange Kabeljaus zu fangen, da diese sich vorzugsweise auf hoher See und damit unerreichbar für ihre damaligen Birkenrindenflöße aufhalten: Der beobachtete Befund unterschätzt also wahrscheinlich noch den Unterschied der durchschnittlichen Größe des Kabeljaus in der Jungsteinzeit, denn die wirklich großen Fische halten sich nicht küstennah, sondern im für die Steinzeitmenschen unerreichbaren Tiefwasser auf. Damals wurden also die größten und ältesten Fische zwangsläufig geschont und dies sicherte Nachwuchs und Durchschnittsgröße der Fänge: Kleine Kabeljauweibchen produzieren nur wenige hunderttausend Eier, große dagegen viele Millionen mit besserer Qualität, so dass die geschlüpften Larven eine größere Überlebenschance haben.
Durch die starke und verbesserte Befischung gerade der fruchtbarsten Altersklassen hat sich die Situation heute sehr verändert. Denn nun werden die großen Exemplare gefangen, während frühreife Kabeljaus begünstigt werden, da sie bereits Nachwuchs produzieren konnten, ehe sie gefangen wurden.
Entsprechendes gilt für das Alter der gefangenen Kabeljaus: Die in der Jungsteinzeit gefangenen Fische waren signifikant älter als die derzeitigen: Mittleres Alter 4,7 Jahre versus 3,1 (1993) bzw. 3,6 (2003).
Außerdem wuchsen junge Kabeljaus in den ersten beiden Lebensjahren signifikant rascher als heute, wo kleine Fische eine größere Chance haben, durch die Fischnetzmaschen zu schlüpfen (kleiner als 35cm 1993, 38cm 2003). Eine biologische Art kann also bezüglich ihrer individuellen Entwicklung rasch auf die Veränderungen ihrer Umwelt reagieren.
[Limburg KE, Walther Y, Hong B, Olson C & Storå J (2008) Prehistoric versus modern Baltic Sea cod fisheries: selectivity across the millennia, Proc. R. Soc. B 275, 2659-2665; Hua Y, Shang H, Tong H, Nehlicha O, Liuc W, Zhaoe C, Yuf J, Wang C, Trinkaus E & Richard MP (2009) Stable isotope dietary analysis of the Tianyuan 1 early modern human, PNAS 106, 10971-10974]
|
 |
| Abb. 1: 37,5% Fitnessabfall pro künstlich aufgezogener Generation. Balken: Standardabweichung. |
|
Tiere in freier Wildbahn unterliegen einem ständigen Selektionsdruck, der unter den vorherrschenden Bedingungen die am wenigsten „fitten“ eliminiert. Kleine Tiere begegnen dem Selektionsdruck gewöhnlich durch eine gewaltige Überproduktion, während bei Großsäugern rituelle Kämpfe um die Weibchen eine große Rolle spielen. Araki et al. (2007) untersuchten den Einfluss der künstlichen Aufzucht von Stahlkopfforellen1 und Lachsen auf die genetische Fitness über mehrere Generationen. Aus künstlichen Brutplätzen für Stahlkopfforellen und Lachse gelangen jedes Jahr mehr als 5 Millionen Jungfische in den Nordpazifik. Dabei fanden die Wissenschaftler heraus, dass die erste Generation in Gefangenschaft aufgezogener Nachkommen von wildlebenden Fischen in 2/3 der Fälle dieselbe biologische Fitness besaß wie nicht in Gefangenschaft aufgewachsene. Aber ab der 2. Generation gab es dann einen raschen konstanten Fitnessabfall bei Fischen, die von Eltern abstammten, die bereits in Gefangenschaft aufgewachsen waren. Je mehr Generationen ihrer Vorfahren bereits in Gefangenschaft aufgewachsen waren, desto größer war der Fitnessabfall (Abb. 1). Die Autoren vermuten, dass die künstliche Aufzucht in gut beschützter – durch Menschen veränderter – Umgebung unbeabsichtigt Zähmung selektiert und die strenge Auswahl der genetisch „besten“ Jungfische lockert.
 |
| Abb. 2: Überleben von Jung- und Erwachsenentieren beider Elefantenarten in Wildnis und Gefangenschaft. |
|
Ein zweites Beispiel, das die Notwendigkeit der Auslese zum Erhalt der Fitness zeigt, kann bei Großsäugern beobachtet werden. Löwen und Elefanten haben ein drastisches Abnehmen in der Wildnis und die große Schwierigkeit, in Gefangenschaft geborene Tiere auszuwildern, gemeinsam. Der Bestand der derzeit noch 23 000 Löwen Afrikas ist bedroht, innerhalb und außerhalb der Nationalparks. Guo (2009) berichtet über den African Lion and Environment Research Trust (ALERT)2, eine 2005 gegründete kommerzielle Organisation, die Tourismus mit der Auswilderung von in Gefangenschaft aufgewachsenen Löwen verbindet. Die Jungtiere werden schrittweise an das Überleben in der Wildnis gewöhnt, nachdem sie im Alter von drei Wochen von ihren Müttern getrennt wurden. Beispielsweise soll das Spazierengehen mit Touristen sie daran gewöhnen, in einem Rudel zu marschieren. Sie werden dann in immer größere Freigehege gesetzt und mit immer mehr Beutetieren statt mit fertiger Nahrung versorgt. Nach Guo sind nur 30% Auswilderungen von Katzenartigen erfolgreich, und dabei handele es sich fast immer um den Transfer von einem natürlichen Lebensraum zu einem anderen. Selbst Zoohaltung ist nicht immer der Garant des Überlebens einer Art, obgleich sie traditionell als solcher angesehen worden ist (Araki 2007): Zoo- und Zirkuspopulationen des afrikanischen (Loxodonta africanus, 300 Tiere in Europa) wie des asiatischen Elefanten (Elephas maximus, 400 Tiere in Europa) reproduzieren sich nicht genug, um ihr Überleben sicherzustellen. Gefangene Elefanten leiden vermehrt an Krankheiten, und die Lebensdauer der erwachsenen Tiere ist gegenüber Wildelefanten deutlich reduziert. Auch hier zeigt sich die Notwendigkeit natürlicher Auslese zum Erhalt der Fitness. Großsäuger in Zoos stellen also wegen der für sie „paradiesischen“ Lebensbedingung ohne Auslese der Bestangepassten kein brauchbares Reservoir für die Wiederbesiedelung der Wildnis dar.
Ein drittes Beispiel: Im Elsass werden der Weißstorch (Ciconia ciconia, Familie Ciconiidae), das Auerhuhn (Tetrao urogallus ssp major, Familie Phasianidae) und der eurasische Luchs (Lynx lynx, Familie Felidae) in den angrenzenden Vogesen wieder eingeführt, mit besserem Erfolg als die afrikanischen Großsäuger, weil vornehmlich wild oder halbwild (im Falle des Storches) aufgewachsene Tiere verwendet werden. In Gefangenschaft aufgewachsene Luchse sind überwiegend unfähig, Beute zu schlagen. Da eine aktuelle Analyse der Weltnaturschutzunion IUCN3 derzeit 17 000 Arten akut vom Aussterben bedroht nennt, darunter jede achte Vogel- und jede vierte Säugetierart, sind „Rettungsversuche“ über Zoos etc. wenig erfolgversprechend. Dass Tiere in Gefangenschaft sehr schnell – binnen historischer, nicht evolutionärer Zeiträume – degenerieren, wusste schon zu Darwins Zeiten Alfred E. Brehm (1829-1884) etwa am Beispiel des Esels, der nur noch ein Schatten seiner wild lebenden Verwandten ist.4 Das von Darwin beschriebene „Survival of the fittest“ ist also ein notwendiger Mechanismus, um Tierpopulationen leistungsfähig zu halten. Dagegen ist bisher ungeklärt, wie aus einer solchen stabilisierenden Selektion echte Neukonstruktion entstehen soll.
[Araki H, Cooper B et al. (2007) Genetic effects of captive breeding cause a rapid, cumulative fitness decline in the wild, Science 218, 100-103; Conolly A (2009), Saving African Lions, Science 325, 146; Clubb R, Rowcliffe M et al. (2008) Compromised survivorship in zoo elephants, Science 322, 1649; Guo J (2009) Will captive breeding save Africa’s king of beasts? Science 324, 331]
Anmerkungen
- 1 Diese Form der Regenbogenforellen (Oncorhynchus mykiss, Familie Salmonidae) wandert zum Laichen einen Arm des Columbia River (nordwestliche USA) flussaufwärts.
- 2 www.lionalert.org
- 3 International Union for the Conservation of Nature
- 4 Brehms Tierleben, Band 7 Säugetiere IV, p. 119ff. Nach der 2. Originalausgabe bearb. von A. Meyer, Gutenberg-Verlag Hamburg 1927.
|
|
Fossilien von Neandertalern und archäologische Untersuchungen an deren Hinterlassenschaften erlauben heute einen durchaus differenzierten Einblick in ihren Körperbau und ihre Lebensweise. Seit den Pionierarbeiten zur Analyse von Nukleinsäuren aus Fossilien liefern auch genetische Studien an fossilen Knochen von Neandertalern Beiträge zu unseren Vorstellungen von diesen Menschen. So legten Krings et al. (1997) erste DNA-Sequenzen aus mitochondrialer DNA (mtDNA) vom Typfossil des Neandertalers vor. Der technische Fortschritt in der Analyse von DNA aus fossilen Proben (Millar et al. 2008) ist auch darin dokumentiert, dass Green et al. (2008) die erste vollständige mitochondriale DNA-Sequenz veröffentlicht haben.
Nach Angaben von A. Briggs hätte die Analyse eines weiteren mitochondrialen Genoms (komplette DNA-Sequenz der Mitochondrien) mit den bisher verfügbaren Technologien ca. 400 Millionen $ erfordert. Er hat jedoch neue Methoden entwickelt, die es erlauben, die mtDNA eines Neandertalers für nur ca. 8000 $ zu sequenzieren (Pennisi 2009). Nun legen Briggs et al. (2009) eine vergleichende Studie an mtDNA von Neandertalern vor, für die fünf vollständige DNA-Sequenzen aus deren Mitochondrien erstellt worden sind.
Ausgangsmaterial für die Studie waren vergleichsweise gut erhaltene Fossilien von Neandertalern aus Russland (Mezmaiskaya-Höhle), Kroatien (Vindija-Höhle), Spanien (El Sidron-Höhle), sowie Fossilien von zwei Individuen aus der kleinen Feldhofer Grotte im Neandertal bei Düsseldorf. Den Fossilien werden Alter von ca. 38 000 bis 70 000 Jahre zugeordnet. Als Referenz diente die mtDNA-Sequenz eines weiteren Neandertalerfossils aus der Vindija-Höhle (Kroatien) (Green et al. 2008).
Die entscheidende technische Entwicklung von Briggs besteht darin, dass er aus einem Extrakt, das aus einer unübersehbar komplexen Mischung verschiedenster DNA-Fragmente (DNA aus Mikroben, von modernen Menschen, DNA aus dem Zellkern) besteht, durch einen molekularbiologischen Trick (Primer extension capture, PEC) die Fragmente aus fossiler mtDNA sehr stark anreichern kann. Mit PEC konnten Proben gewonnen werden, die zu 18,2 – 40,2% aus mitochondrialer DNA bestehen (dies entspricht einer Anreichung um das 3640 – 80400 fache ).
Ein Vergleich der mitochondrialen Genome weist bei 16 565 Basenpaaren (bp) 55 variable Positionen auf. Durchschnittlich unterscheiden sich die sechs mtDNA der Neandertaler um 20,2 bp voneinander. Im Vergleich dazu weichen die mitochondrialen Genome moderner Menschen um das Dreifache voneinander ab, wenn man von global verteilten Proben ausgeht. Verglichen mit modernen Europäern sind die individuellen Abweichungen der mtDNA-Sequenzen der Neandertaler um 37% geringer.
Briggs et al. versuchten mit statistischen Methoden die „effektive Populationsgröße“ der Neandertaler zu ermitteln. Diese kann man bei Zugrundelegung bestimmter Modelle anhand der Variation von DNA-Sequenzen bestimmen. So ergibt sich für Schweden bei einer Bevölkerung von ca. 9 Millionen eine effektive Population (der Anteil, der zur Fortpflanzung beiträgt) von 100 000. Die effektive Populationsgröße der Neandertaler wurde mit 3500 an der Fortpflanzung beteiligten weiblichen Neandertalerinnen (oder 7000 weiblichen und männlichen Individuen insgesamt) bestimmt. Das lässt auf eine wirkliche Populationsgröße von ca. 70 000 Neandertalern schließen, was einer vergleichsweise kleinen Population entspricht. Diese eher geringe Größe der Population von Neandertalern könnte für ihr Aussterben von Bedeutung gewesen sein.
Phylogenetische Analysen der neuen Daten bestätigen bisherige Vorstellungen zur Ähnlichkeit von Neandertalern mit modernen Menschen und anderen Primaten. Die mtDNA-Daten separieren die Neandertaler von modernen Menschen – und zwar sowohl von den Afrikanern als auch von den Nicht-Afrikanern, gleichzeitig sind aber Neandertaler und moderne Menschen gemeinsam klar von anderen Primaten (Schimpanse und Bonobo) getrennt. Die bisher vorliegenden Studien basieren auf vergleichsweise wenigen Genomdaten. Sie bedürfen einer Bestätigung durch weitere Untersuchungen. Die dazu nötigen Methoden und Techniken liegen inzwischen vor, wie Briggs et al. (2009) eindrucksvoll zeigen, und so darf man gespannt sein auf die bereits angekündigten Arbeiten über DNA-Sequenzen aus dem Zellkern von Neandertalern.
[Briggs AW, Good JM et al. (2009) Targeted retrieval and analysis of five Neandertal mtDNA genomes. Science 325, 318-321; Green RE et al. (2008) A complete Neanderthal mitochondrial genome sequence determinated by high-throughout sequencing. Cell 134, 416-426; Krings M, Stone A, Schmitz RW, Krainitzki H & Pääbo S (1997) Neandertal DNA sequence and the origin of modern humans. Cell 90, 19-30; Millar CD, Huynen L, Subramanian S, Mohandesan E & Lambert DM (2008) New developements in ancient genomics. Trends Ecol. Evol. 23, 386-393; Pennisi E (2009) Sequencing Neandertal mitochondrial genomes by the half-dozen. Science 325, 252.]
|
|
 |

